

Engineered Fluorescent Strains of Cryptococcus neoformans: a Versatile Toolbox for Studies of Host-Pathogen Interactions and Fungal Biology, Including the Viable but Nonculturable State
- PMID: 36005449
- PMCID: PMC9603711
- DOI: 10.1128/spectrum.01504-22
Engineered Fluorescent Strains of Cryptococcus neoformans: a Versatile Toolbox for Studies of Host-Pathogen Interactions and Fungal Biology, Including the Viable but Nonculturable State
Abstract
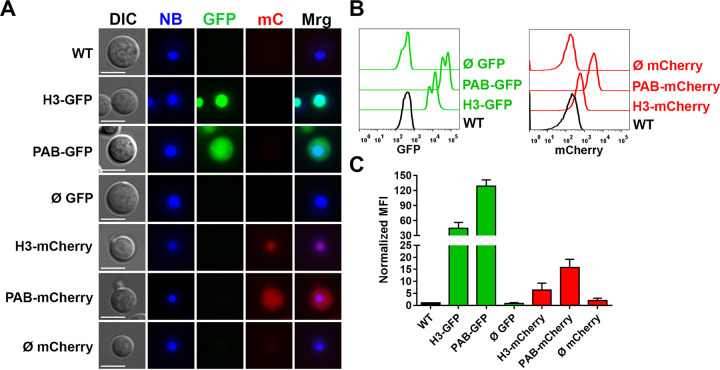
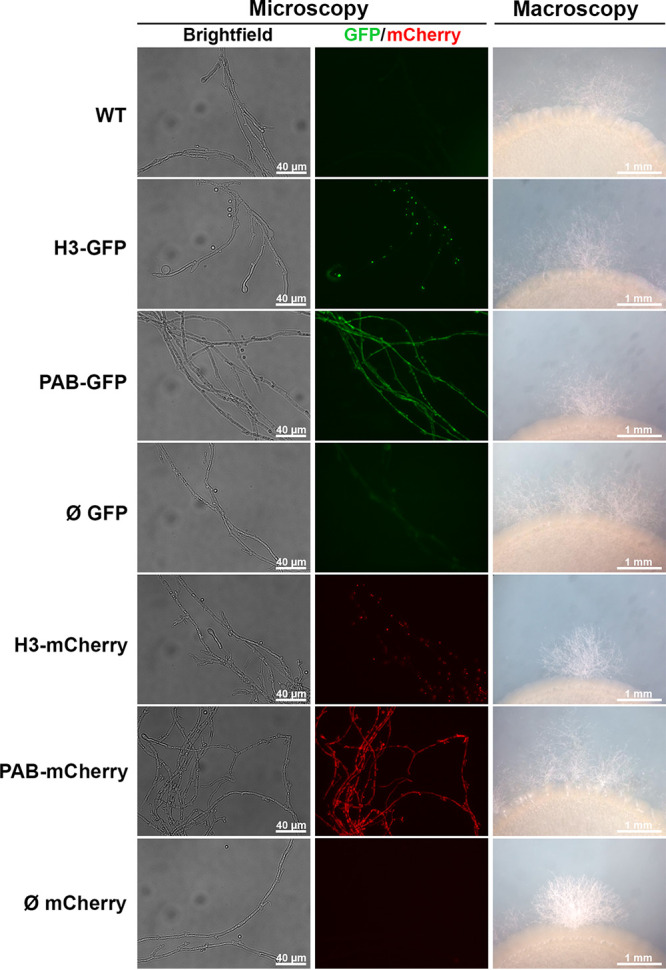
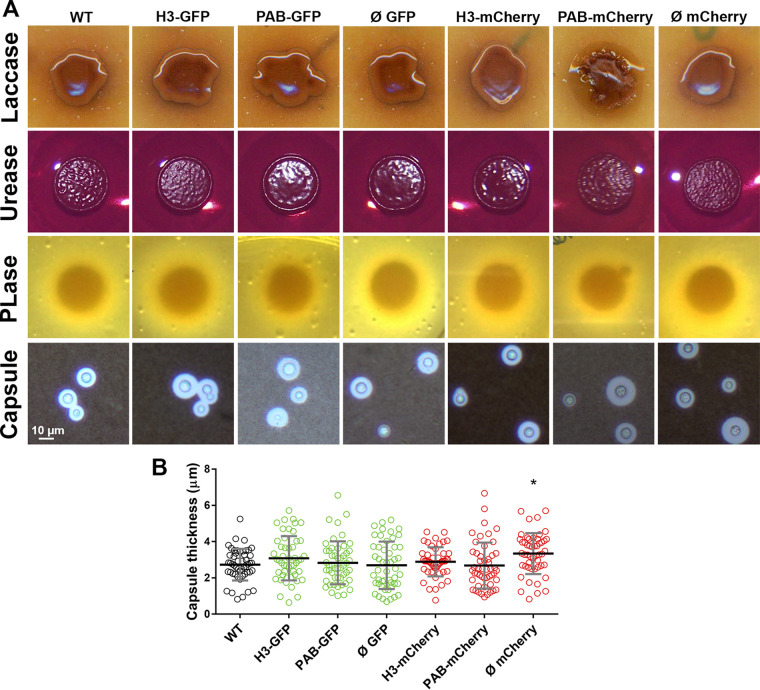
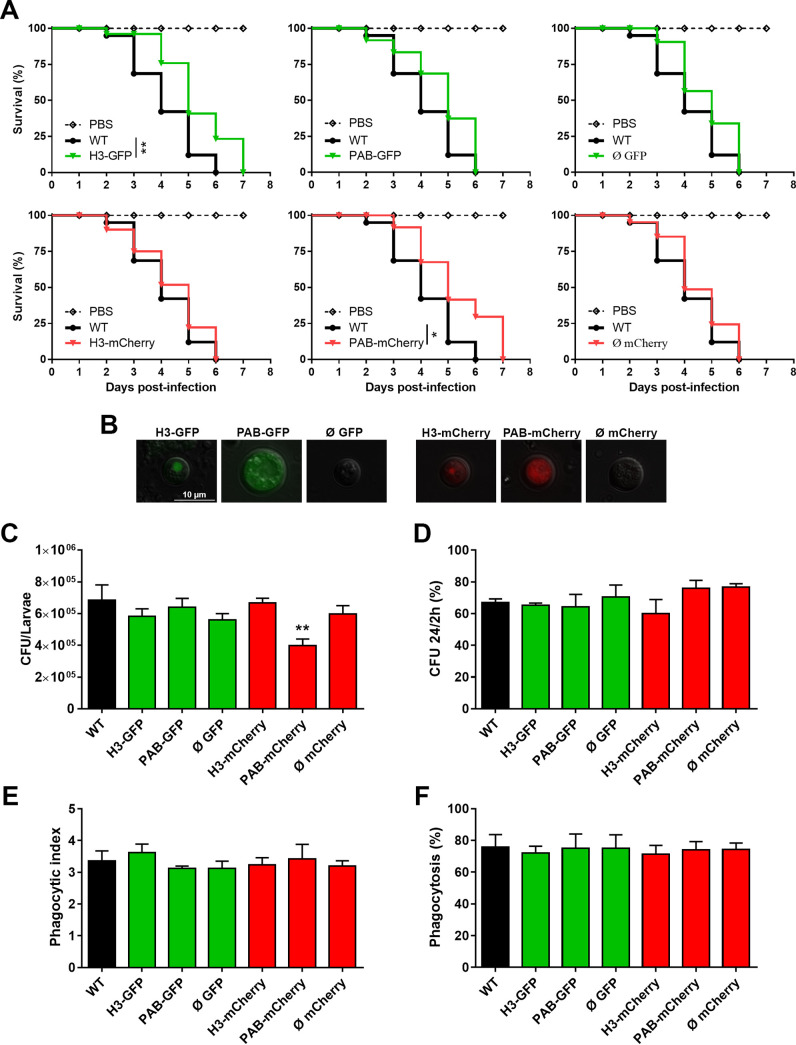
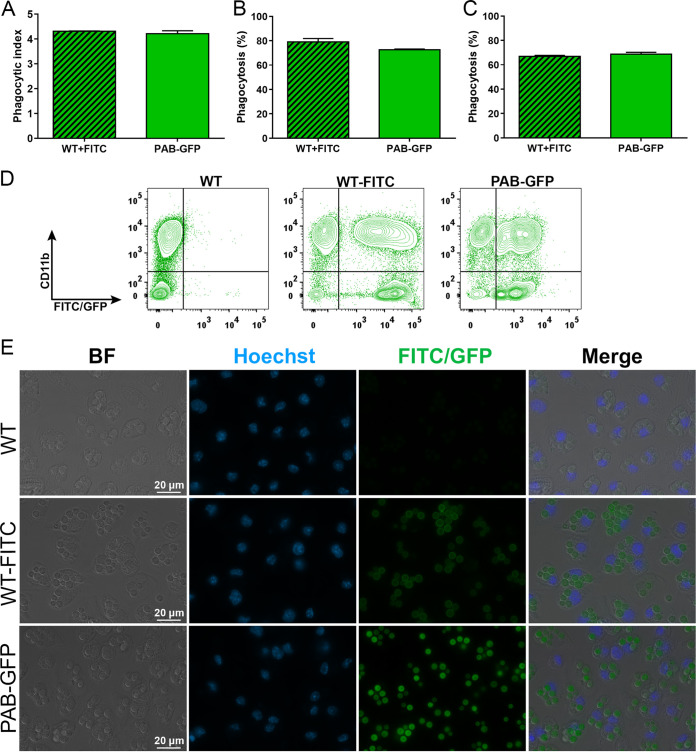
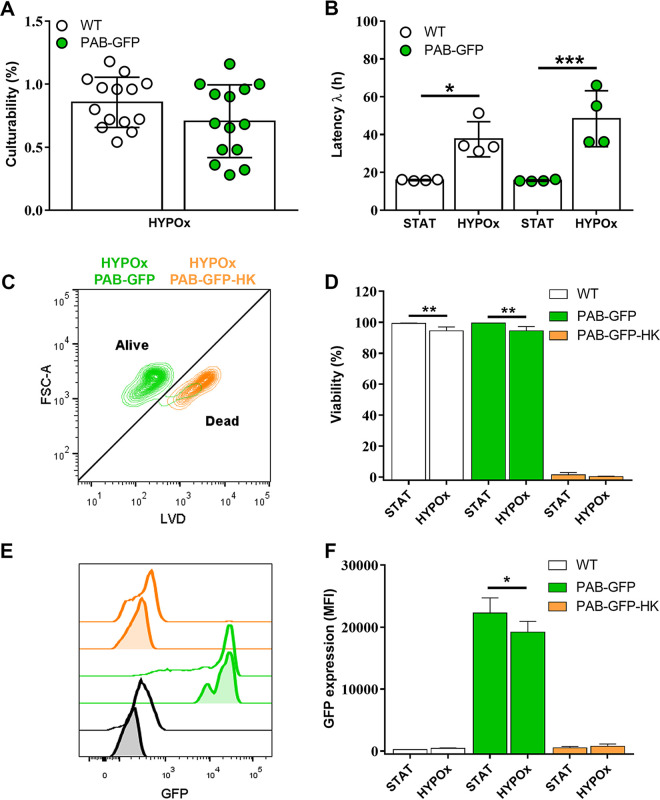
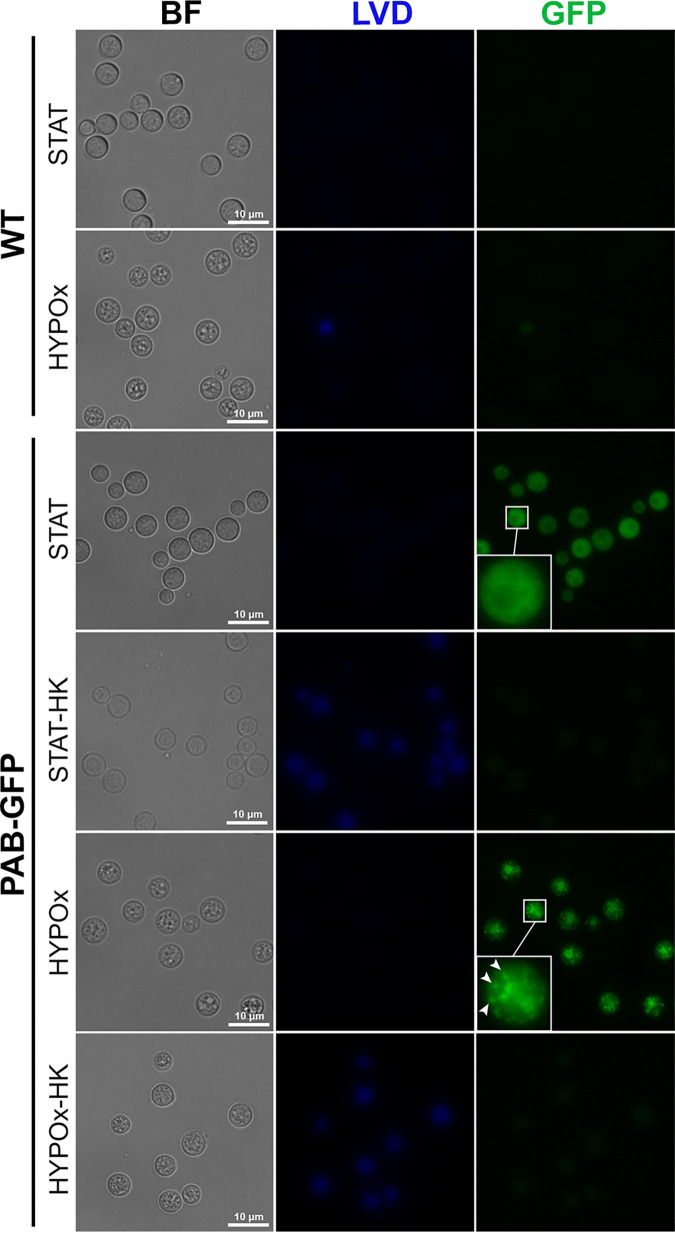
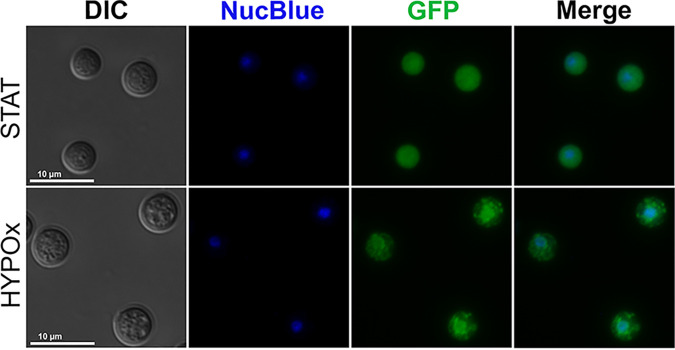
Cryptococcus neoformans is an opportunistic fungal pathogen known for its remarkable ability to infect and subvert phagocytes. This ability provides survival and persistence within the host and relies on phenotypic plasticity. The viable but nonculturable (VBNC) phenotype was recently described in C. neoformans, whose study is promising in understanding the pathophysiology of cryptococcosis. The use of fluorescent strains is improving host interaction research, but it is still underexploited. Here, we fused histone H3 or the poly(A) binding protein (Pab) to enhanced green fluorescent protein (eGFP) or mCherry, obtaining a set of C. neoformans transformants with different colors, patterns of fluorescence, and selective markers (hygromycin B resistance [Hygr] or neomycin resistance [Neor]). We validated their similarity to the parental strain in the stress response, the expression of virulence-related phenotypes, mating, virulence in Galleria mellonella, and survival within murine macrophages. PAB-GFP, the brightest transformant, was successfully applied for the analysis of phagocytosis by flow cytometry and fluorescence microscopy. Moreover, we demonstrated that an engineered fluorescent strain of C. neoformans was able to generate VBNC cells. GFP-tagged Pab1, a key regulator of the stress response, evidenced nuclear retention of Pab1 and the assembly of cytoplasmic stress granules, unveiling posttranscriptional mechanisms associated with dormant C. neoformans cells. Our results support that the PAB-GFP strain is a useful tool for research on C. neoformans. IMPORTANCE Cryptococcus neoformans is a human-pathogenic yeast that can undergo a dormant state and is responsible for over 180,000 deaths annually worldwide. We engineered a set of fluorescent transformants to aid in research on C. neoformans. A mutant with GFP-tagged Pab1 improved fluorescence-based techniques used in host interaction studies. Moreover, this mutant induced a viable but nonculturable phenotype and uncovered posttranscriptional mechanisms associated with dormant C. neoformans. The experimental use of fluorescent mutants may shed light on C. neoformans-host interactions and fungal biology, including dormant phenotypes.
Keywords: Cryptococcus neoformans; GFP; H99; VBNC; dormancy; fluorescently tagged strains; histone H3; mCherry; poly(A) binding protein; stress granules.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Kicking sleepers out of bed: Macrophages promote reactivation of dormant Cryptococcus neoformans by extracellular vesicle release and non-lytic exocytosis.PLoS Pathog. 2023 Nov 30;19(11):e1011841. doi: 10.1371/journal.ppat.1011841. eCollection 2023 Nov. PLoS Pathog. 2023. PMID: 38033163 Free PMC article.
-
Cryptococcus neoformans host adaptation: toward biological evidence of dormancy.mBio. 2015 Mar 31;6(2):e02580-14. doi: 10.1128/mBio.02580-14. mBio. 2015. PMID: 25827423 Free PMC article.
-
Identification of Aph1, a phosphate-regulated, secreted, and vacuolar acid phosphatase in Cryptococcus neoformans.mBio. 2014 Sep 16;5(5):e01649-14. doi: 10.1128/mBio.01649-14. mBio. 2014. PMID: 25227465 Free PMC article.
-
Dormancy in Cryptococcus neoformans: 60 years of accumulating evidence.J Clin Invest. 2020 Jul 1;130(7):3353-3360. doi: 10.1172/JCI136223. J Clin Invest. 2020. PMID: 32484459 Free PMC article. Review.
-
The interplay of phenotype and genotype in Cryptococcus neoformans disease.Biosci Rep. 2020 Oct 30;40(10):BSR20190337. doi: 10.1042/BSR20190337. Biosci Rep. 2020. PMID: 33021310 Free PMC article. Review.
Cited by
-
Kicking sleepers out of bed: Macrophages promote reactivation of dormant Cryptococcus neoformans by extracellular vesicle release and non-lytic exocytosis.PLoS Pathog. 2023 Nov 30;19(11):e1011841. doi: 10.1371/journal.ppat.1011841. eCollection 2023 Nov. PLoS Pathog. 2023. PMID: 38033163 Free PMC article.
-
Induction of Dormancy in Cryptococcus neoformans In Vitro: The HypNOS Protocol.Methods Mol Biol. 2024;2775:349-358. doi: 10.1007/978-1-0716-3722-7_22. Methods Mol Biol. 2024. PMID: 38758328
-
Population heterogeneity in Cryptococcus neoformans: Impact on pathogenesis.PLoS Pathog. 2024 Jul 11;20(7):e1012332. doi: 10.1371/journal.ppat.1012332. eCollection 2024 Jul. PLoS Pathog. 2024. PMID: 38990818 Free PMC article. Review. No abstract available.
-
Is Cryptococcus neoformans a pleomorphic fungus?Curr Opin Microbiol. 2024 Dec;82:102539. doi: 10.1016/j.mib.2024.102539. Epub 2024 Sep 10. Curr Opin Microbiol. 2024. PMID: 39260180 Free PMC article. Review.
References
-
- Rhodes J, Desjardins CA, Sykes SM, Beale MA, Vanhove M, Sakthikumar S, Chen Y, Gujja S, Saif S, Chowdhary A, Lawson DJ, Ponzio V, Colombo AL, Meyer W, Engelthaler DM, Hagen F, Illnait-Zaragozi MT, Alanio A, Vreulink J-M, Heitman J, Perfect JR, Litvintseva AP, Bicanic T, Harrison TS, Fisher MC, Cuomo CA. 2017. Tracing genetic exchange and biogeography of Cryptococcus neoformans var. grubii at the global population level. Genetics 207:327–346. doi:10.1534/genetics.117.203836. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
