

Composition and organization of kinetochores show plasticity in apicomplexan chromosome segregation
- PMID: 36006241
- PMCID: PMC9418836
- DOI: 10.1083/jcb.202111084
Composition and organization of kinetochores show plasticity in apicomplexan chromosome segregation
Abstract
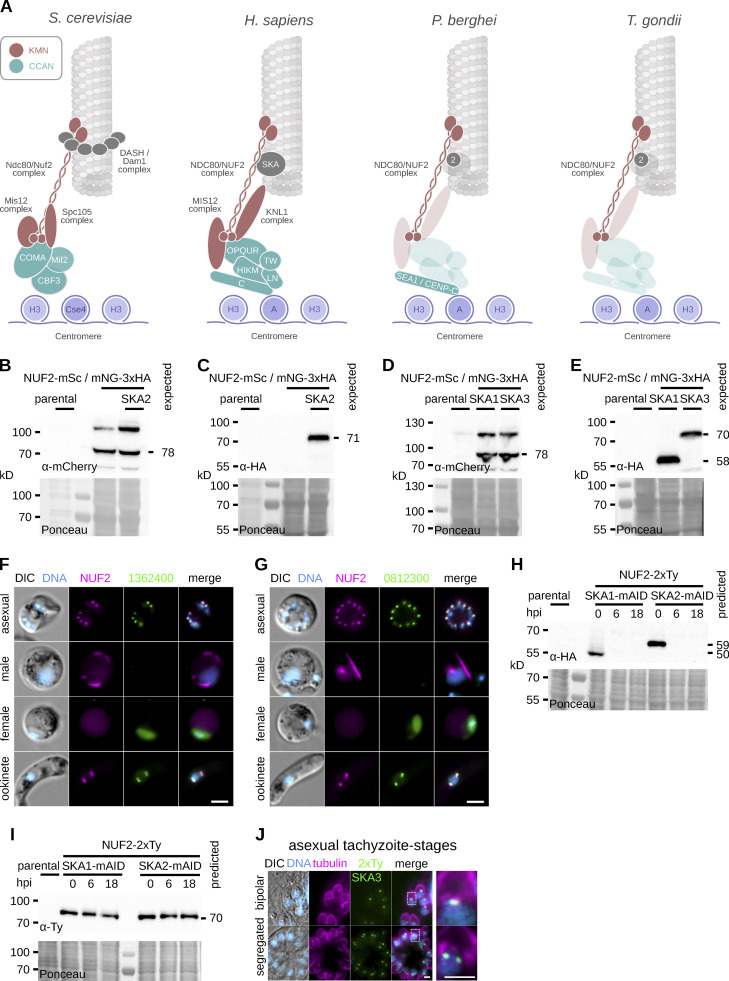
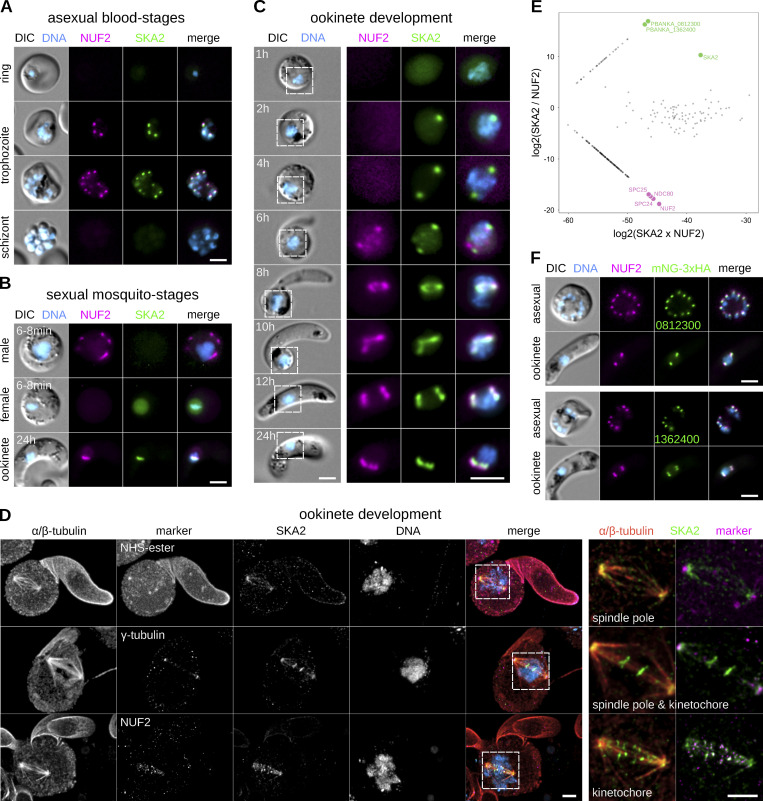
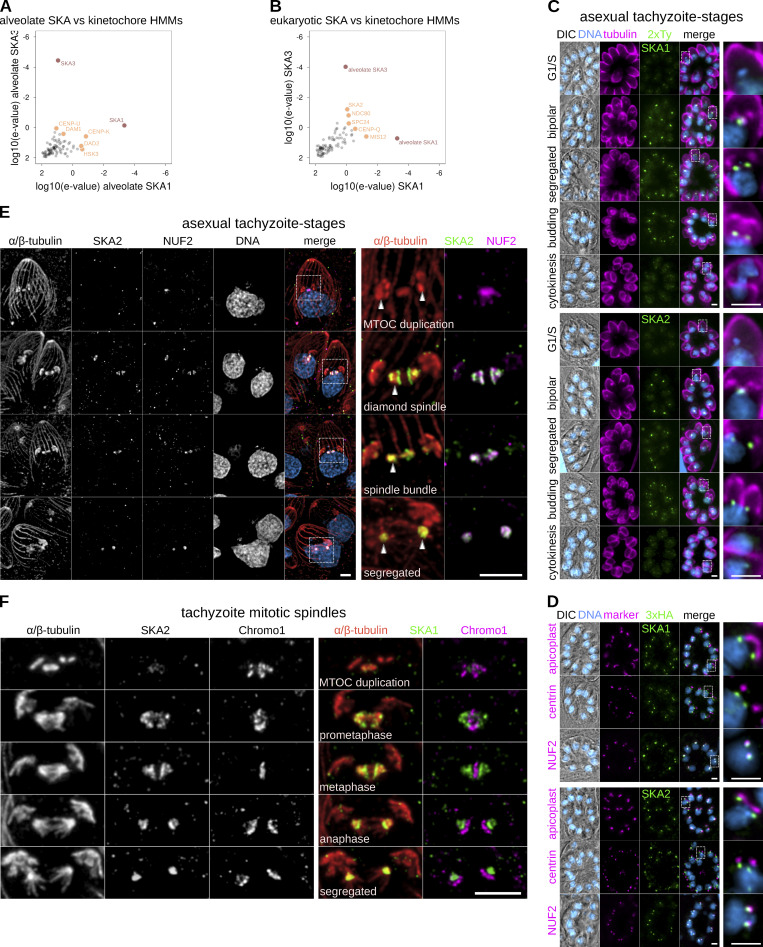
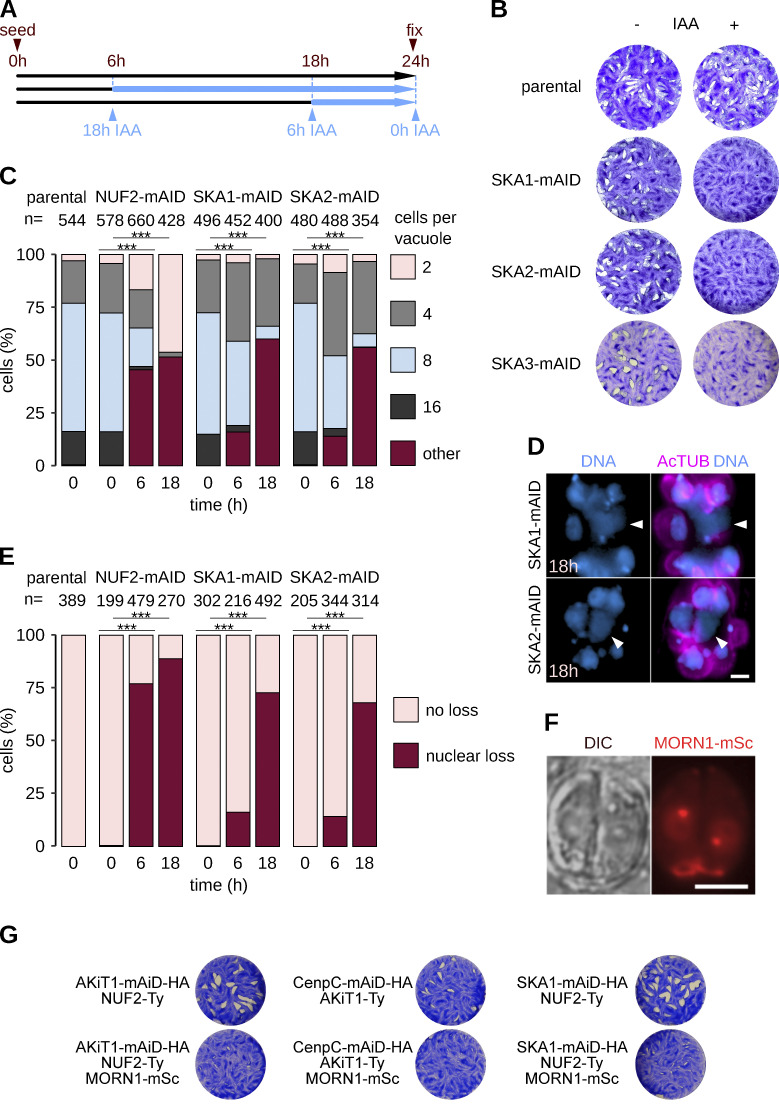
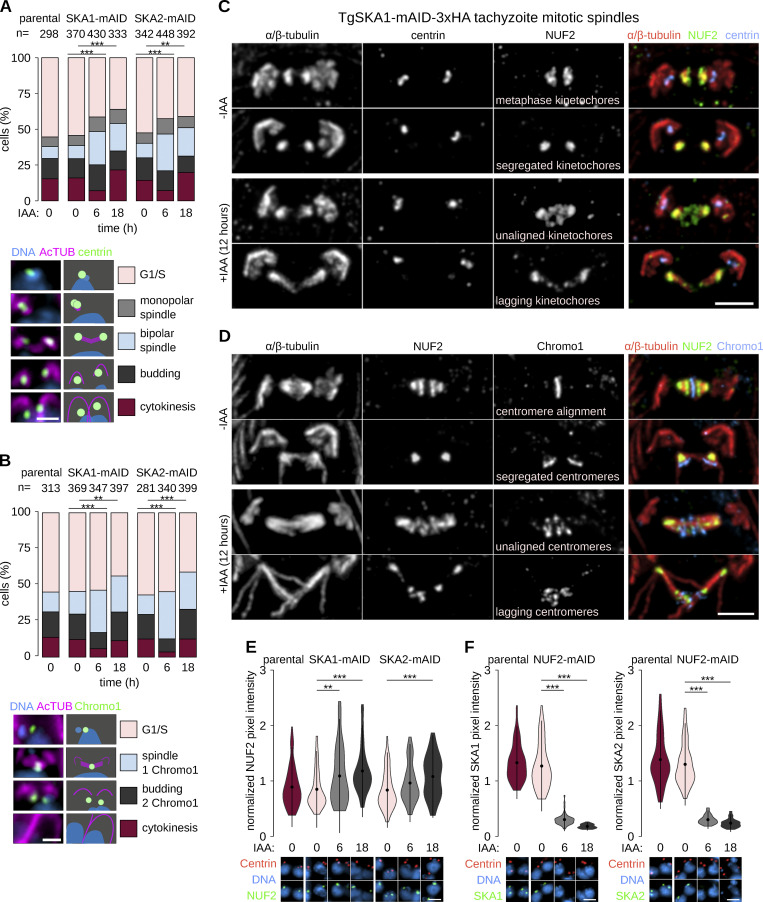
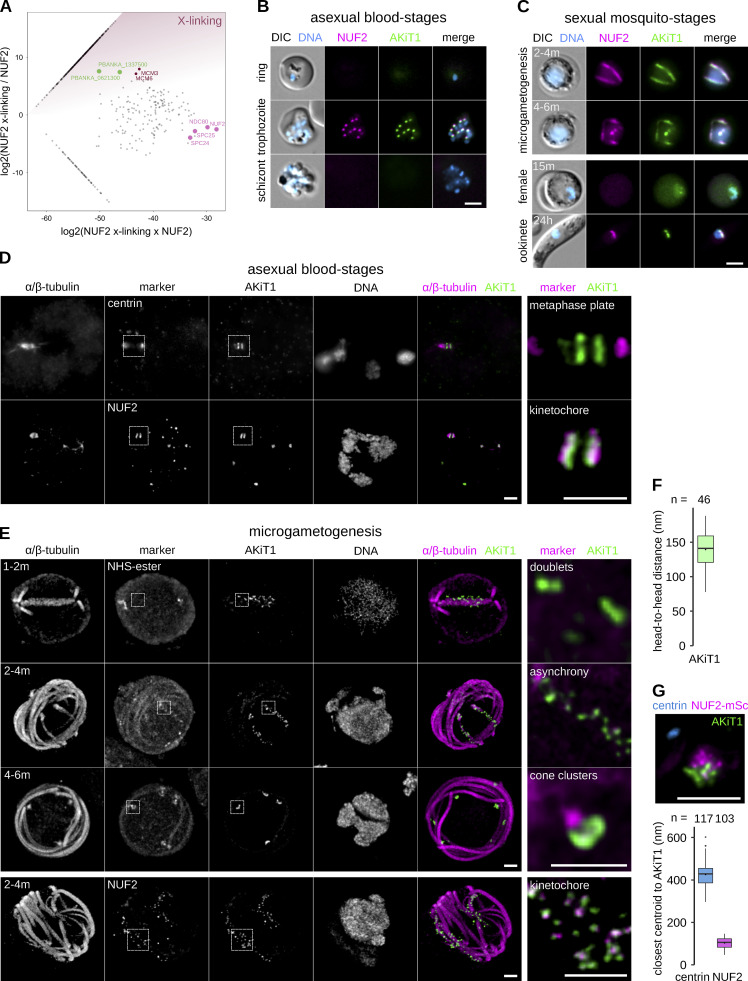
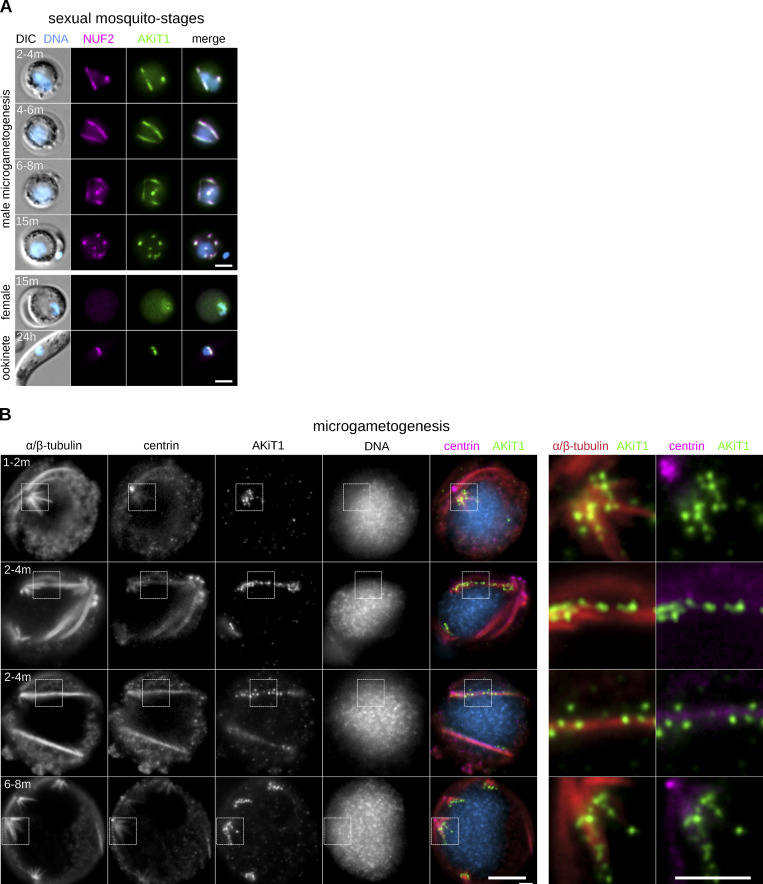
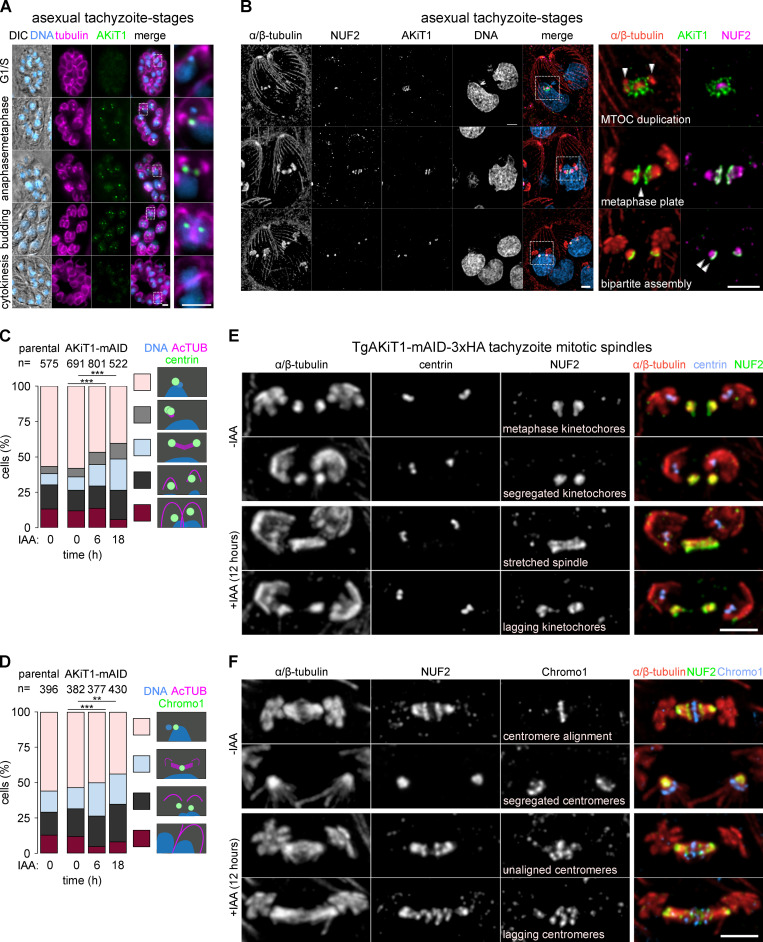
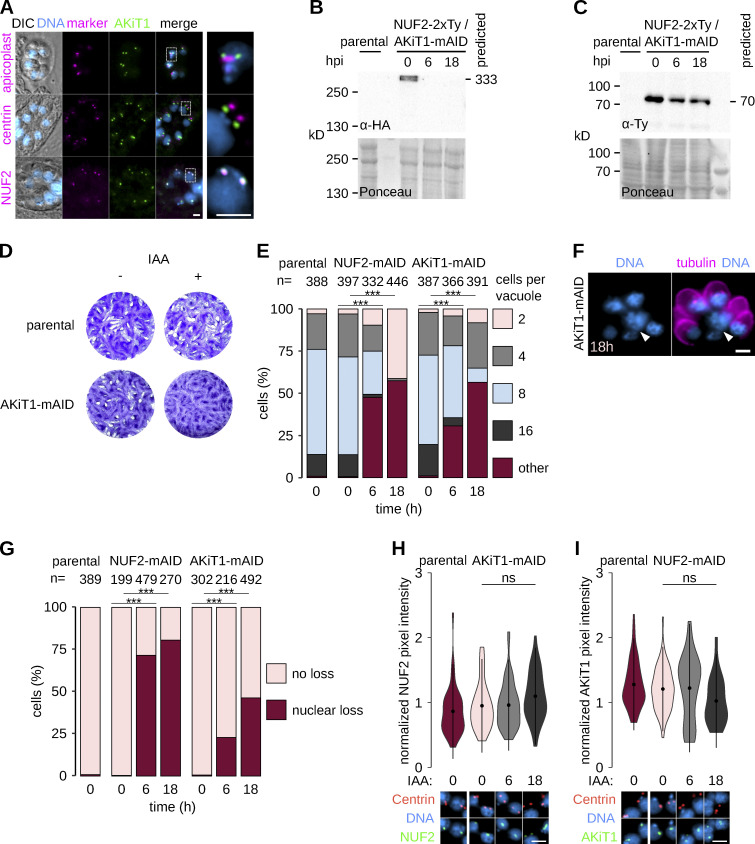
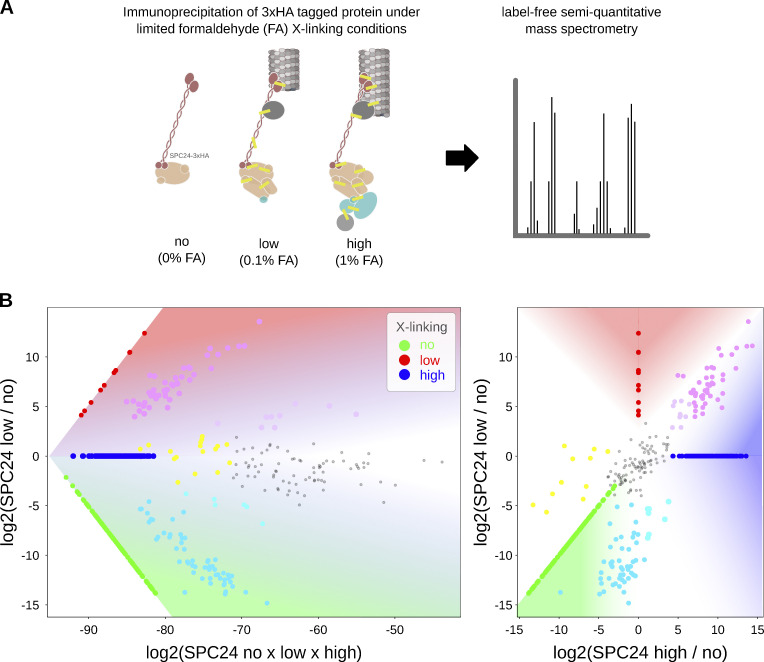
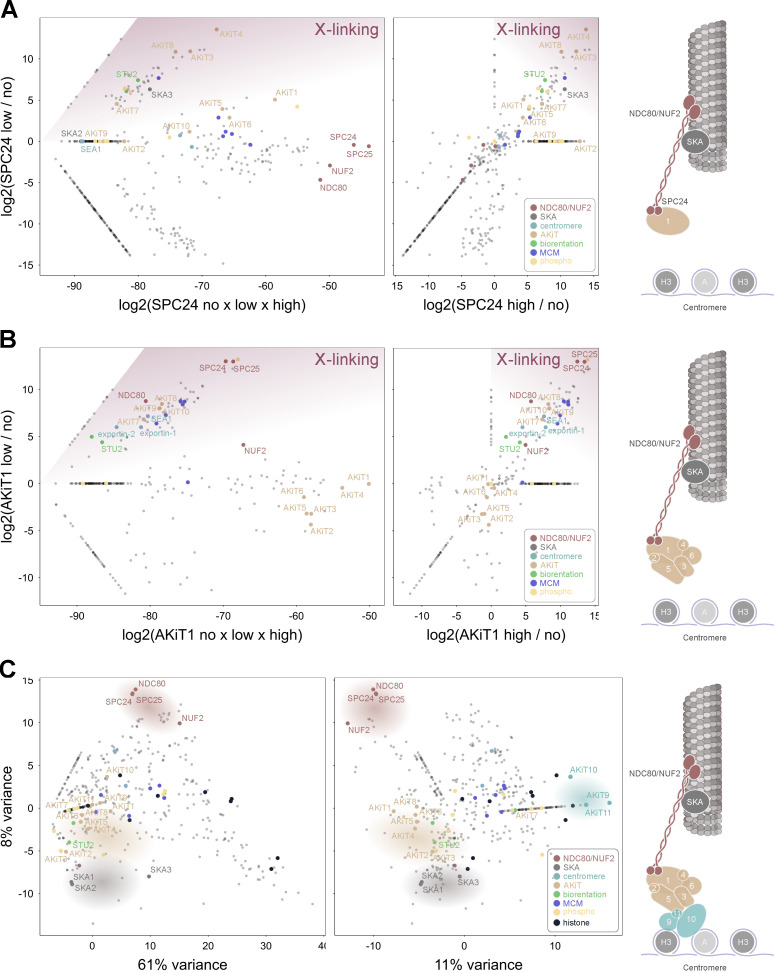
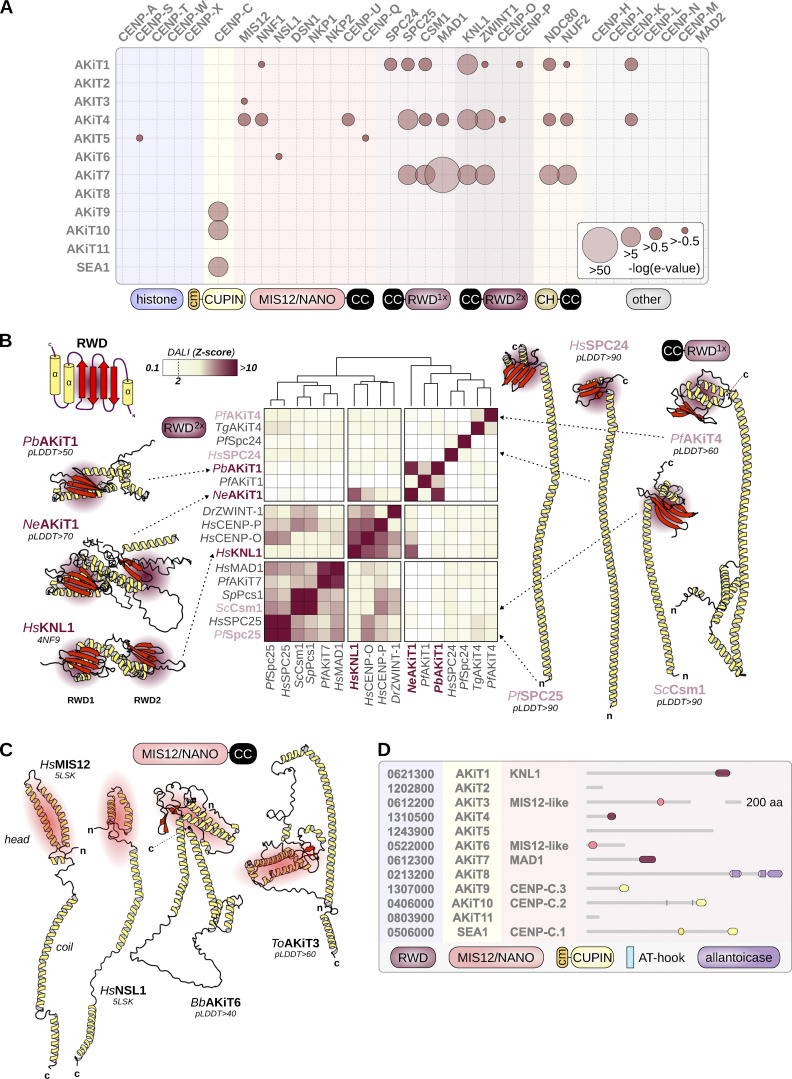
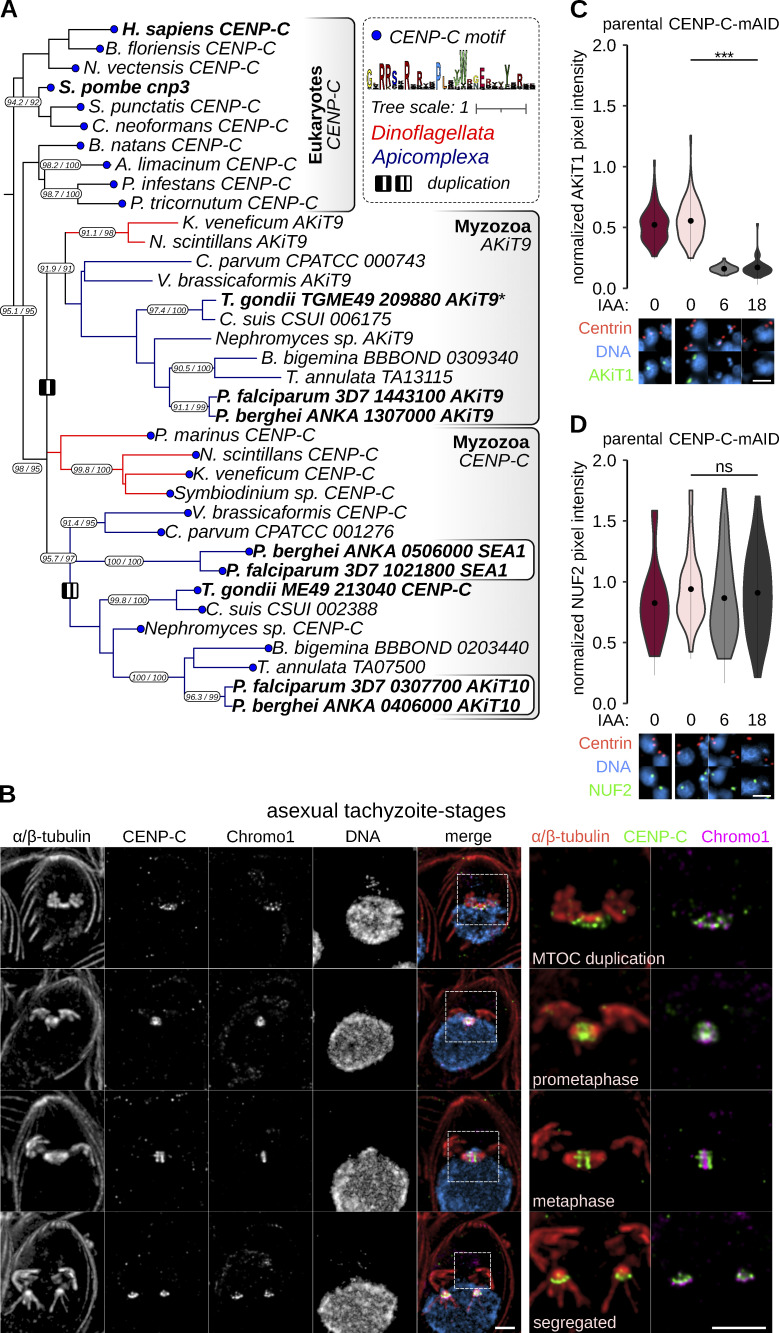
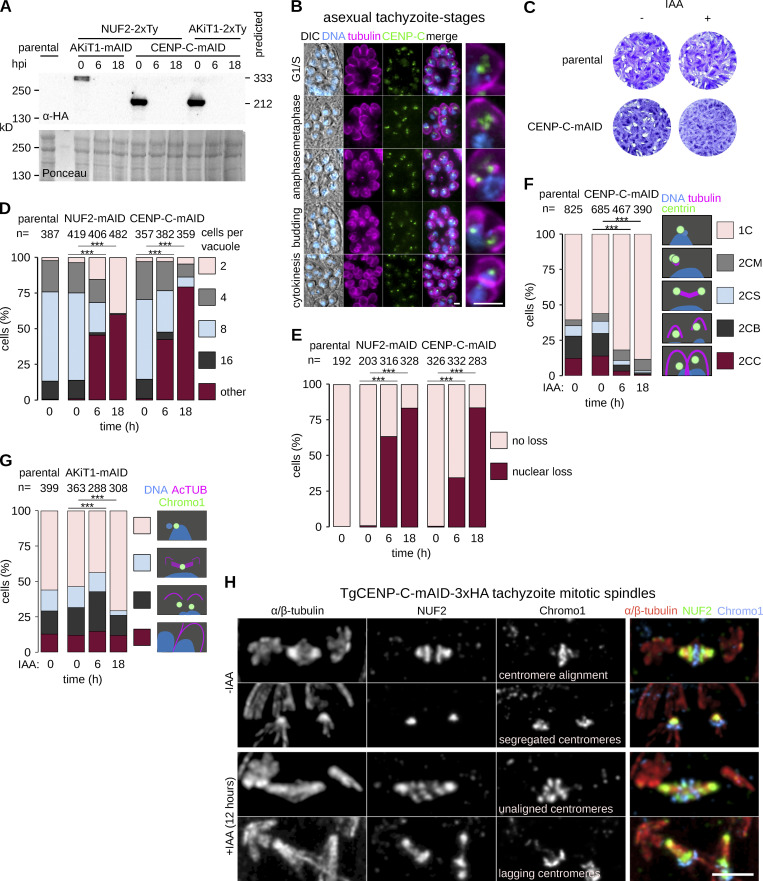
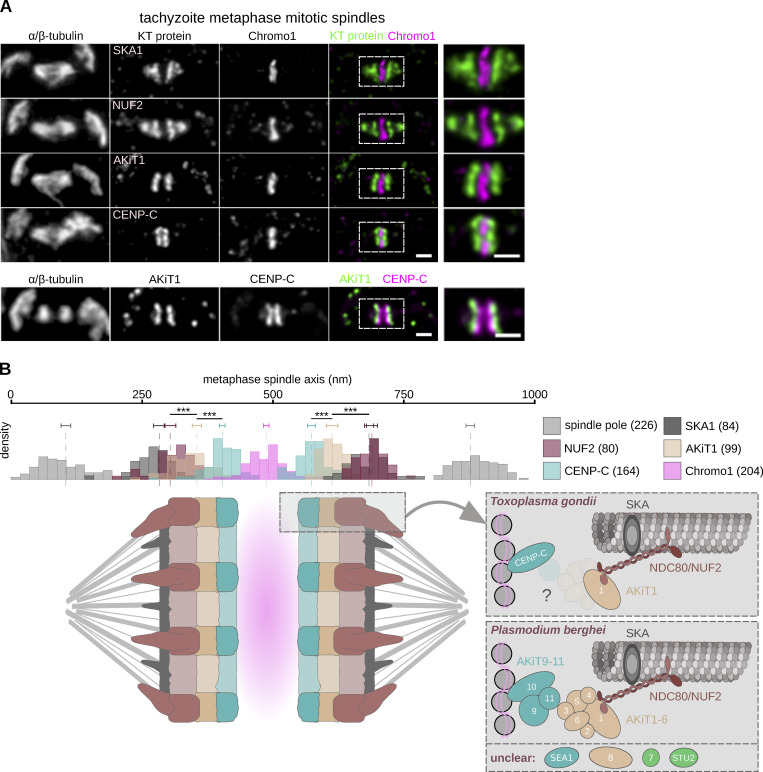
Kinetochores are multiprotein assemblies directing mitotic spindle attachment and chromosome segregation. In apicomplexan parasites, most known kinetochore components and associated regulators are apparently missing, suggesting a minimal structure with limited control over chromosome segregation. In this study, we use interactomics combined with deep homology searches to identify 13 previously unknown components of kinetochores in Apicomplexa. Apicomplexan kinetochores are highly divergent in sequence and composition from animal and fungal models. The nanoscale organization includes at least four discrete compartments, each displaying different biochemical interactions, subkinetochore localizations and evolutionary rates across the phylum. We reveal alignment of kinetochores at the metaphase plate in both Plasmodium berghei and Toxoplasma gondii, suggestive of a conserved "hold signal" that prevents precocious entry into anaphase. Finally, we show unexpected plasticity in kinetochore composition and segregation between apicomplexan lifecycle stages, suggestive of diverse requirements to maintain fidelity of chromosome segregation across parasite modes of division.
© 2022 Brusini et al.
Figures



Similar articles
-
Real-time dynamics of Plasmodium NDC80 reveals unusual modes of chromosome segregation during parasite proliferation.J Cell Sci. 2020 Jun 30;134(5):jcs245753. doi: 10.1242/jcs.245753. J Cell Sci. 2020. PMID: 32501284 Free PMC article.
-
Mechanisms of chromosome biorientation and bipolar spindle assembly analyzed by computational modeling.Elife. 2020 Feb 13;9:e48787. doi: 10.7554/eLife.48787. Elife. 2020. PMID: 32053104 Free PMC article.
-
Sad1 Spatiotemporally Regulates Kinetochore Clustering To Ensure High-Fidelity Chromosome Segregation in the Human Fungal Pathogen Cryptococcus neoformans.mSphere. 2018 Jul 5;3(4):e00190-18. doi: 10.1128/mSphere.00190-18. mSphere. 2018. PMID: 29976642 Free PMC article.
-
Merotelic kinetochores in mammalian tissue cells.Philos Trans R Soc Lond B Biol Sci. 2005 Mar 29;360(1455):553-68. doi: 10.1098/rstb.2004.1610. Philos Trans R Soc Lond B Biol Sci. 2005. PMID: 15897180 Free PMC article. Review.
-
Monitoring the fidelity of mitotic chromosome segregation by the spindle assembly checkpoint.Cell Prolif. 2011 Oct;44(5):391-400. doi: 10.1111/j.1365-2184.2011.00767.x. Cell Prolif. 2011. PMID: 21951282 Free PMC article. Review.
Cited by
-
Divergent Plasmodium kinases drive MTOC, kinetochore and axoneme organisation in male gametogenesis.Life Sci Alliance. 2025 Mar 24;8(6):e202403056. doi: 10.26508/lsa.202403056. Print 2025 Jun. Life Sci Alliance. 2025. PMID: 40127922 Free PMC article.
-
A novel SUN1-ALLAN complex coordinates segregation of the bipartite MTOC across the nuclear envelope during rapid closed mitosis in Plasmodium berghei.Elife. 2025 May 20;14:RP106537. doi: 10.7554/eLife.106537. Elife. 2025. PMID: 40392232 Free PMC article.
-
Enzymatically enhanced ultrastructure expansion microscopy unlocks expansion of in vitro Toxoplasma gondii cysts.mSphere. 2024 Sep 25;9(9):e0032224. doi: 10.1128/msphere.00322-24. Epub 2024 Aug 27. mSphere. 2024. PMID: 39189782 Free PMC article.
-
Apicomplexan mitoribosome from highly fragmented rRNAs to a functional machine.Nat Commun. 2024 Dec 17;15(1):10689. doi: 10.1038/s41467-024-55033-z. Nat Commun. 2024. PMID: 39690155 Free PMC article.
-
An atypical Arp2/3 complex is required for Plasmodium DNA segregation and malaria transmission.Nat Microbiol. 2025 Jul;10(7):1775-1790. doi: 10.1038/s41564-025-02023-6. Epub 2025 Jun 13. Nat Microbiol. 2025. PMID: 40514561 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
