

Hand2os1 Regulates the Secretion of Progesterone in Mice Corpus Luteum
- PMID: 36006319
- PMCID: PMC9415164
- DOI: 10.3390/vetsci9080404
Hand2os1 Regulates the Secretion of Progesterone in Mice Corpus Luteum
Abstract
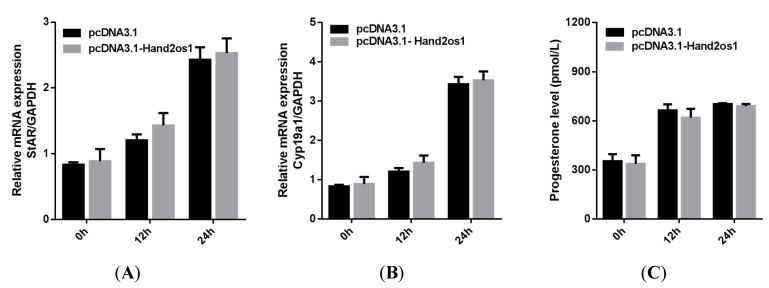
The corpus luteum plays a key role in pregnancy maintenance and estrous cycle regulation by secreting progesterone. Hand2os1 is an lncRNA located upstream of Hand2, with which a bidirectional promoter is shared and is involved in the regulation of cardiac development and embryo implantation in mice. The aim of this study was to investigate the expression and regulation of Hand2os1 in the ovaries. Here, we used RNAscope to detect differential expression of Hand2os1 in the ovaries of cycling and pregnant mice. Hand2os1 was specifically detected in luteal cells during the proestrus and estrus phases, showing its highest expression in the corpus luteum at estrus. Additionally, Hand2os1 was strongly expressed in the corpus luteum on day 4 of pregnancy, but the positive signal progressively disappeared after day 8, was detected again on day 18, and gradually decreased after delivery. Hand2os1 significantly promoted the synthesis of progesterone and the expression of StAR and Cyp11a1. The decreased progesterone levels caused by Hand2os1 interference were rescued by the overexpression of StAR. Our findings suggest that Hand2os1 may regulate the secretion of progesterone in the mouse corpus luteum by affecting the key rate-limiting enzyme StAR, which may have an impact on the maintenance of pregnancy.
Keywords: Hand2os1; corpus luteum; lncRNA; mouse; progesterone.
Conflict of interest statement
The authors declare that there is no conflict of interest that could be perceived as prejudicing the impartiality of the research reported.
Figures






Similar articles
-
The Role of Reduced Oxygen Supply and Transcription Factors cJUN and CREB in Progesterone Production during the Corpus Luteum Rescue in Gilts.Animals (Basel). 2022 Oct 20;12(20):2855. doi: 10.3390/ani12202855. Animals (Basel). 2022. PMID: 36290240 Free PMC article.
-
Progesterone-induced RNA Hand2os1 regulates decidualization in mice uteri.Reproduction. 2020 Mar;159(3):303-314. doi: 10.1530/REP-19-0401. Reproduction. 2020. PMID: 31990672
-
The lncRNA Hand2os1/Uph locus orchestrates heart development through regulation of precise expression of Hand2.Development. 2019 Jul 4;146(13):dev176198. doi: 10.1242/dev.176198. Development. 2019. PMID: 31273086
-
Utero-ovarian relationships in placental mammals: role of uterus and embryo in the regulation of progesterone secretion by the corpus luteum. A review.Reprod Nutr Dev (1980). 1983;23(5):793-816. doi: 10.1051/rnd:19830601. Reprod Nutr Dev (1980). 1983. PMID: 6359305 Review.
-
Mechanisms controlling the function and life span of the corpus luteum.Physiol Rev. 2000 Jan;80(1):1-29. doi: 10.1152/physrev.2000.80.1.1. Physiol Rev. 2000. PMID: 10617764 Review.
Cited by
-
LncRNA NEAT1 participates in diminished ovarian reserve by affecting granulosa cell apoptosis and estradiol synthesis via the miR-204-5p/ESR1 axis.J Ovarian Res. 2025 May 16;18(1):102. doi: 10.1186/s13048-025-01683-6. J Ovarian Res. 2025. PMID: 40380255 Free PMC article.
-
Structural analysis and core promoter prediction of STAR gene and its regulatory mechanism of progesterone synthesis in bovine luteal cells.Sci Rep. 2025 Mar 5;15(1):7746. doi: 10.1038/s41598-025-92446-2. Sci Rep. 2025. PMID: 40044781 Free PMC article.
References
-
- Proietto S., Cortasa S.A., Corso M.C., Inserra P.I.F., Charif S.E., Schmidt A.R., Di Giorgio N.P., Lux-Lantos V., Vitullo A.D., Dorfman V.B., et al. Prolactin Is a Strong Candidate for the Regulation of Luteal Steroidogenesis in Vizcachas (Lagostomus maximus) Int. J. Endocrinol. 2018;2018:1910672. doi: 10.1155/2018/1910672. - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
