

Lysosomal GPCR-like protein LYCHOS signals cholesterol sufficiency to mTORC1
- PMID: 36007018
- PMCID: PMC10023259
- DOI: 10.1126/science.abg6621
Lysosomal GPCR-like protein LYCHOS signals cholesterol sufficiency to mTORC1
Abstract
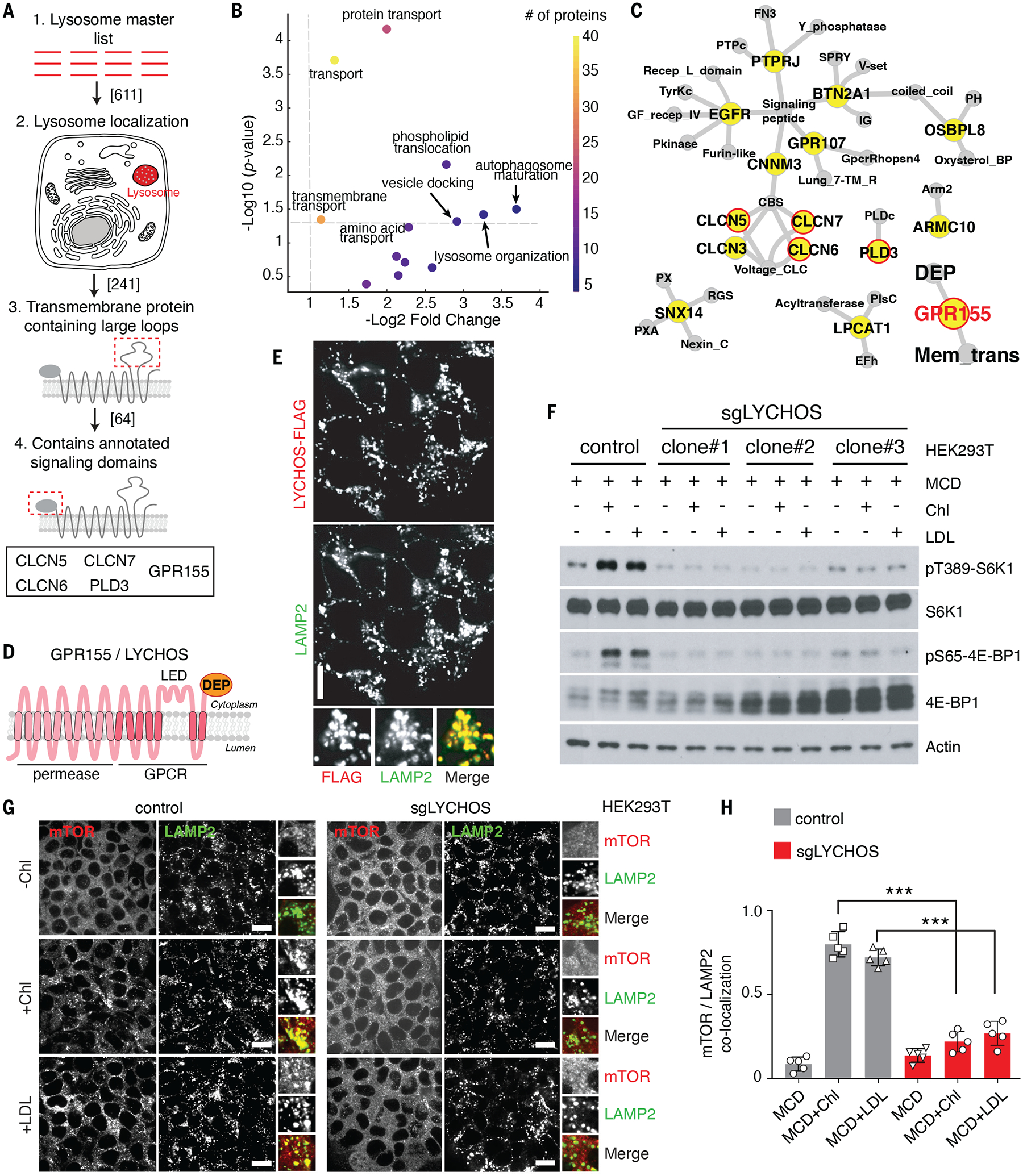
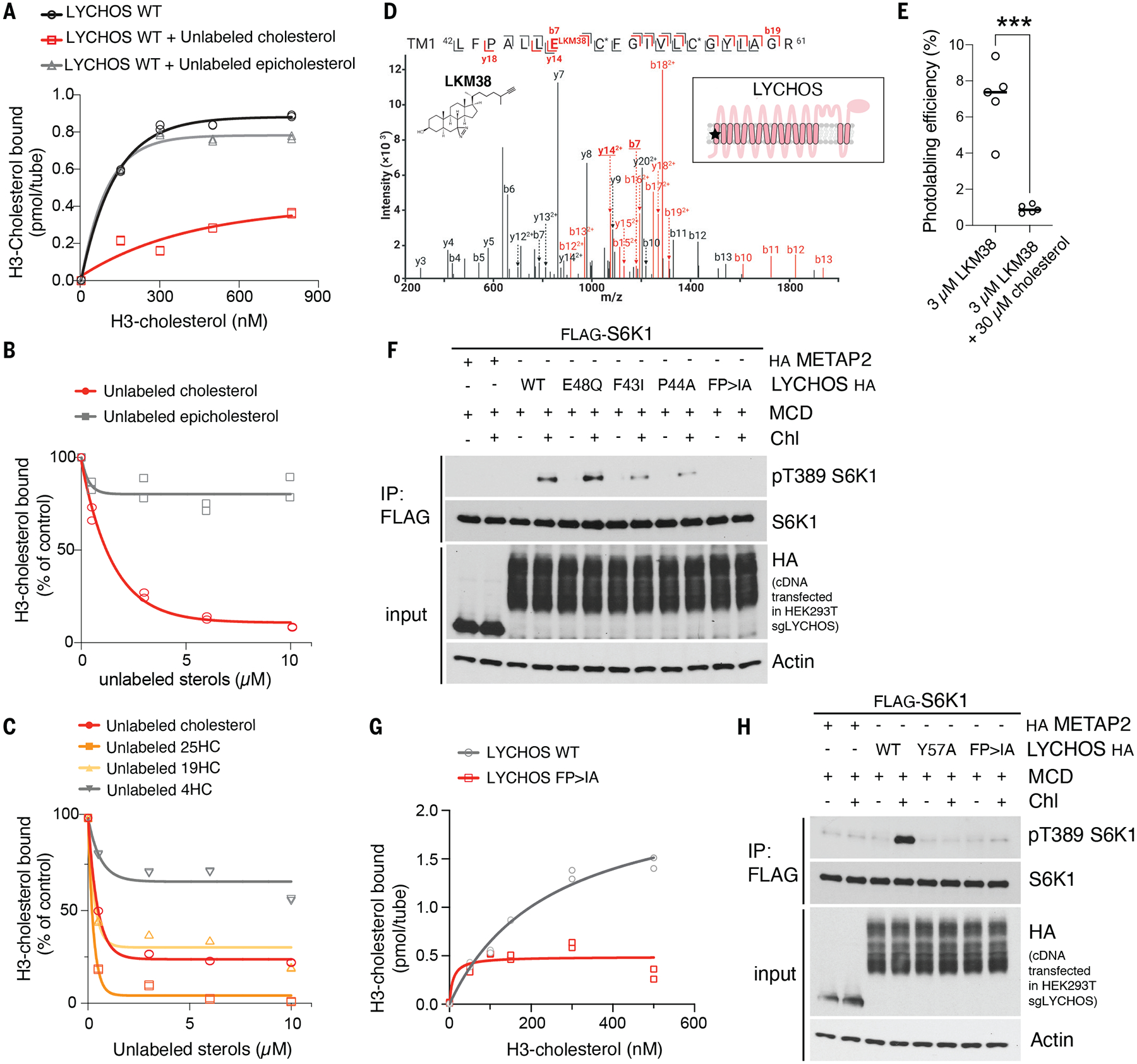
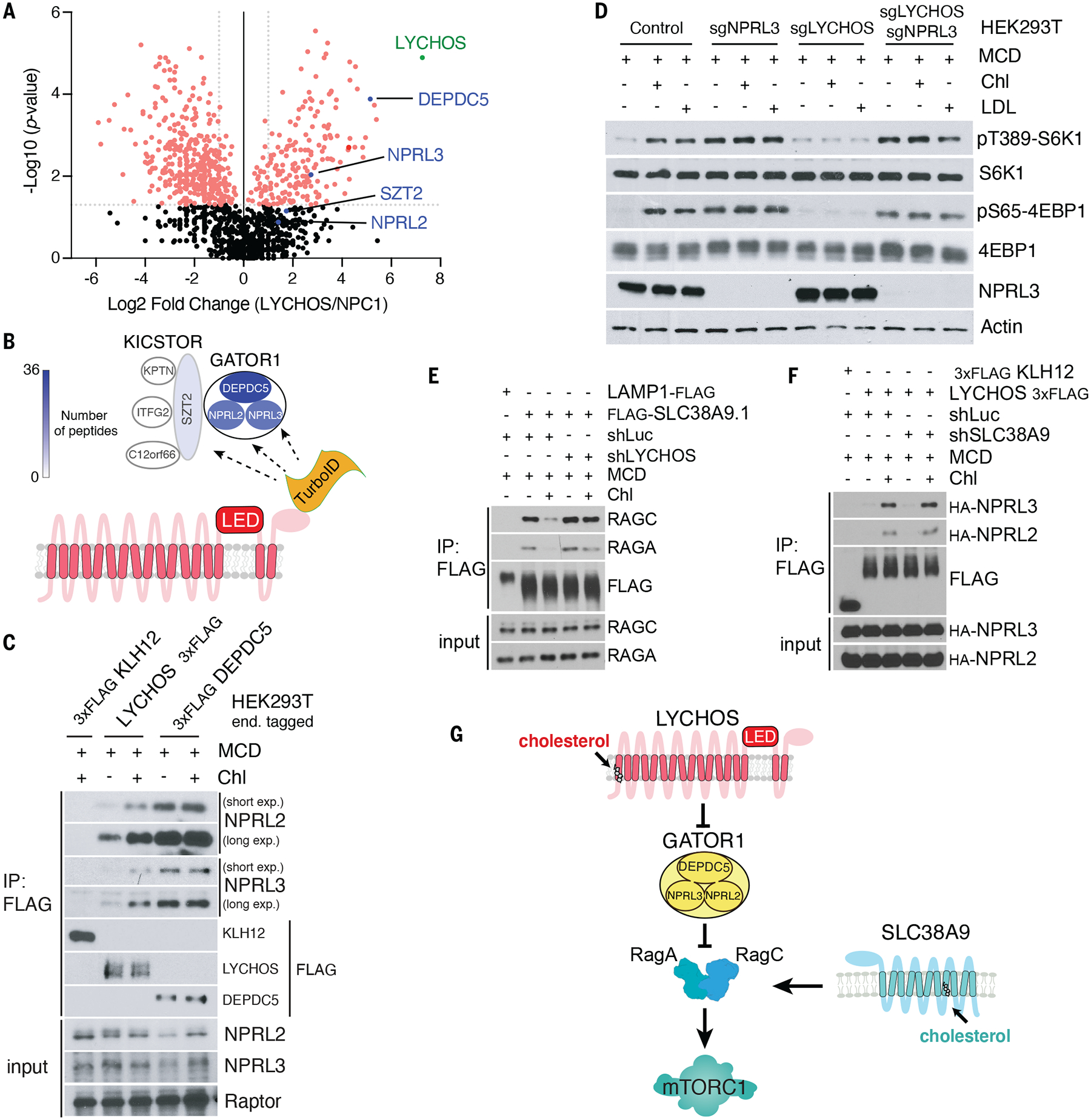
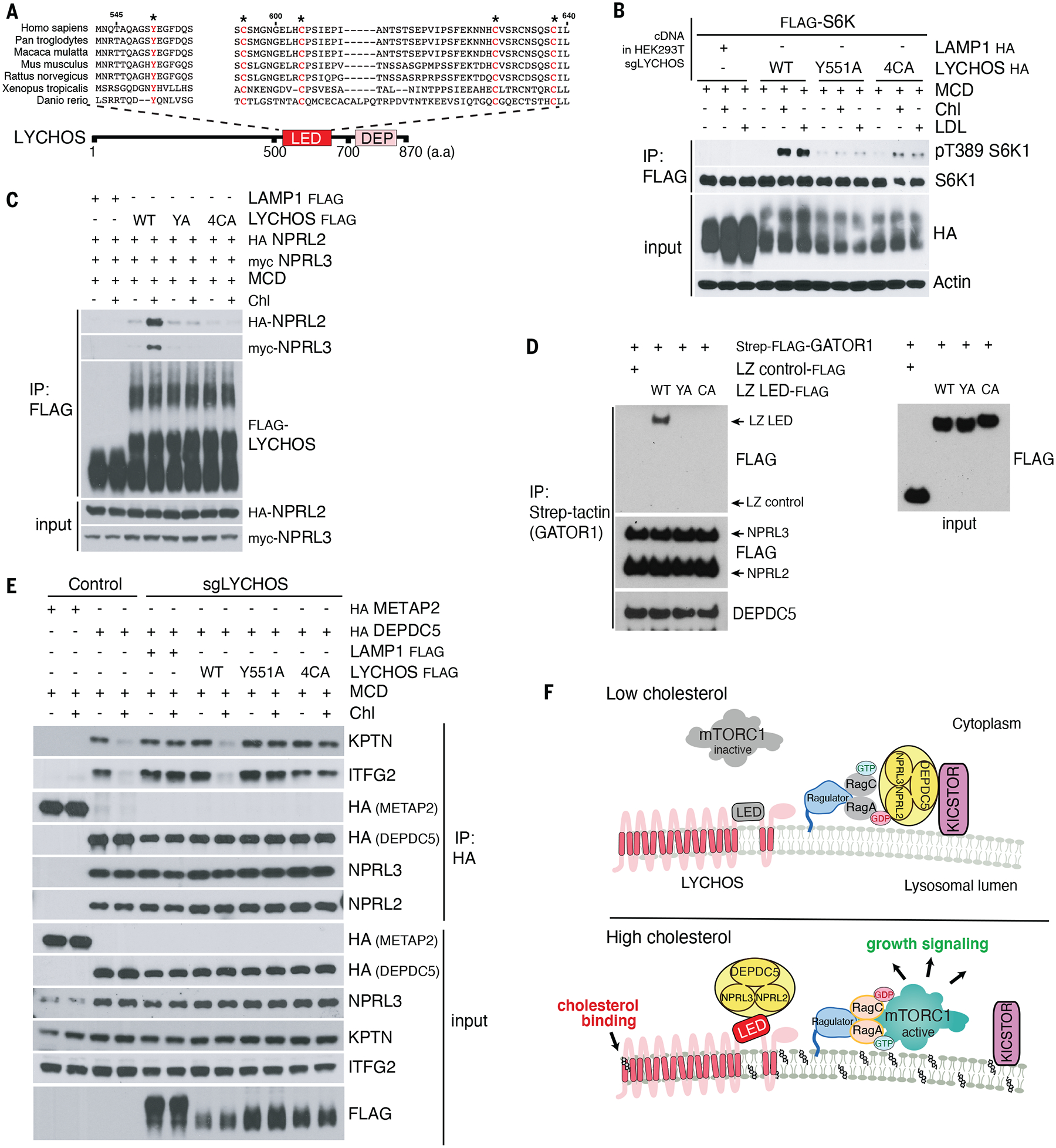
Lysosomes coordinate cellular metabolism and growth upon sensing of essential nutrients, including cholesterol. Through bioinformatic analysis of lysosomal proteomes, we identified lysosomal cholesterol signaling (LYCHOS, previously annotated as G protein-coupled receptor 155), a multidomain transmembrane protein that enables cholesterol-dependent activation of the master growth regulator, the protein kinase mechanistic target of rapamycin complex 1 (mTORC1). Cholesterol bound to the amino-terminal permease-like region of LYCHOS, and mutating this site impaired mTORC1 activation. At high cholesterol concentrations, LYCHOS bound to the GATOR1 complex, a guanosine triphosphatase (GTPase)-activating protein for the Rag GTPases, through a conserved cytoplasm-facing loop. By sequestering GATOR1, LYCHOS promotes cholesterol- and Rag-dependent recruitment of mTORC1 to lysosomes. Thus, LYCHOS functions in a lysosomal pathway for cholesterol sensing and couples cholesterol concentrations to mTORC1-dependent anabolic signaling.
Conflict of interest statement
Figures
Comment in
-
mTOR gets greasy: lysosomal sensing of cholesterol.Cell Res. 2023 Mar;33(3):189-190. doi: 10.1038/s41422-022-00740-9. Cell Res. 2023. PMID: 36284233 Free PMC article. No abstract available.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
