

The convergent xenogeneic silencer MucR predisposes α-proteobacteria to integrate AT-rich symbiosis genes
- PMID: 36007892
- PMCID: PMC9410896
- DOI: 10.1093/nar/gkac664
The convergent xenogeneic silencer MucR predisposes α-proteobacteria to integrate AT-rich symbiosis genes
Abstract
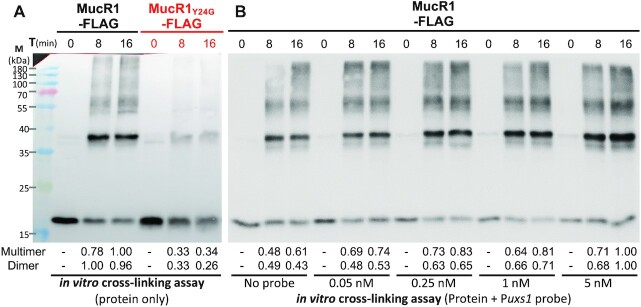
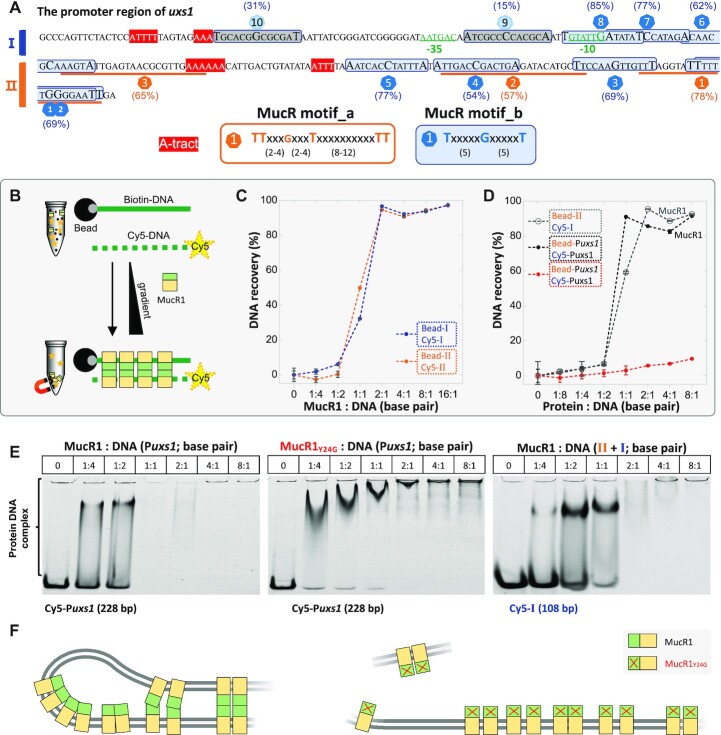
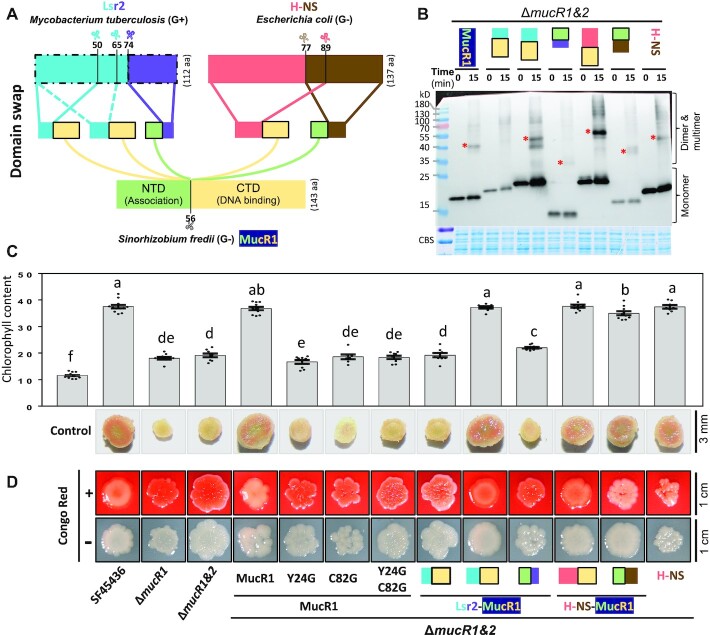
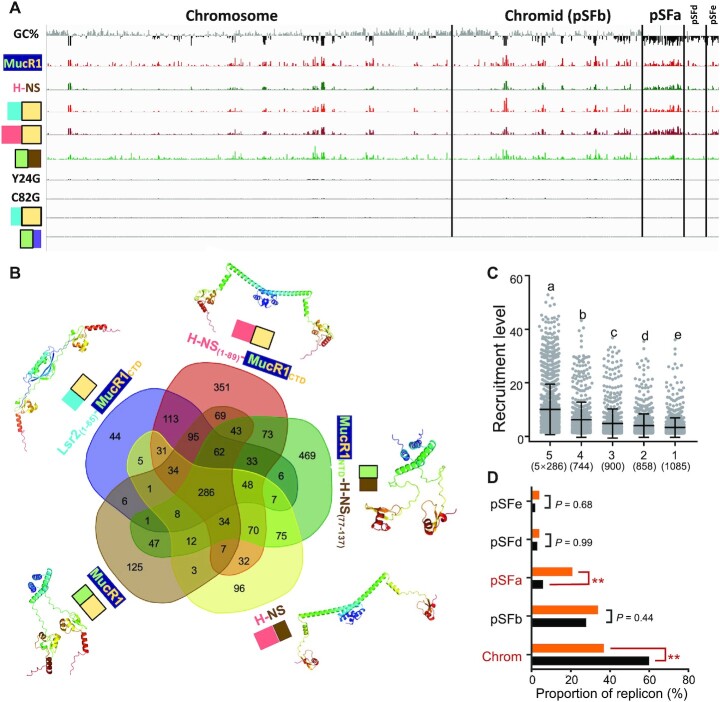
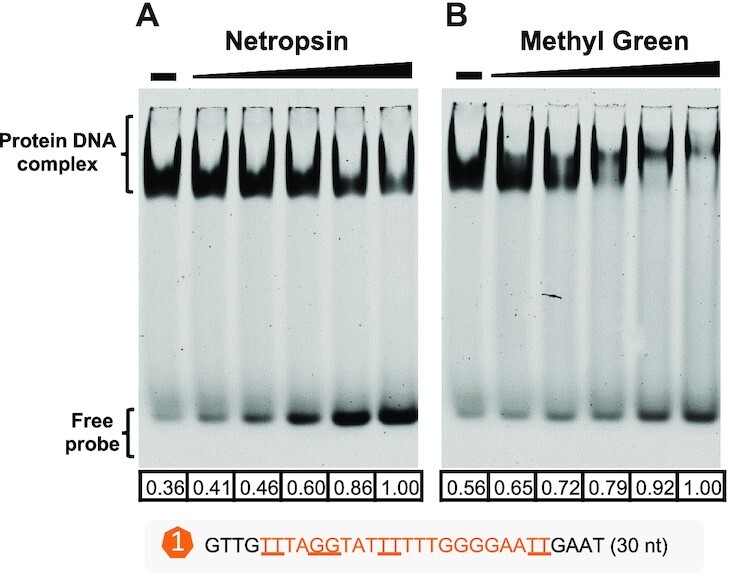
Bacterial adaptation is largely shaped by horizontal gene transfer, xenogeneic silencing mediated by lineage-specific DNA bridgers (H-NS, Lsr2, MvaT and Rok), and various anti-silencing mechanisms. No xenogeneic silencing DNA bridger is known for α-proteobacteria, from which mitochondria evolved. By investigating α-proteobacterium Sinorhizobium fredii, a facultative legume microsymbiont, here we report the conserved zinc-finger bearing MucR as a novel xenogeneic silencing DNA bridger. Self-association mediated by its N-terminal domain (NTD) is required for DNA-MucR-DNA bridging complex formation, maximizing MucR stability, transcriptional silencing, and efficient symbiosis in legume nodules. Essential roles of NTD, CTD (C-terminal DNA-binding domain), or full-length MucR in symbiosis can be replaced by non-homologous NTD, CTD, or full-length protein of H-NS from γ-proteobacterium Escherichia coli, while NTD rather than CTD of Lsr2 from Gram-positive Mycobacterium tuberculosis can replace the corresponding domain of MucR in symbiosis. Chromatin immunoprecipitation sequencing reveals similar recruitment profiles of H-NS, MucR and various functional chimeric xenogeneic silencers across the multipartite genome of S. fredii, i.e. preferring AT-rich genomic islands and symbiosis plasmid with key symbiosis genes as shared targets. Collectively, the convergently evolved DNA bridger MucR predisposed α-proteobacteria to integrate AT-rich foreign DNA including symbiosis genes, horizontal transfer of which is strongly selected in nature.
© The Author(s) 2022. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
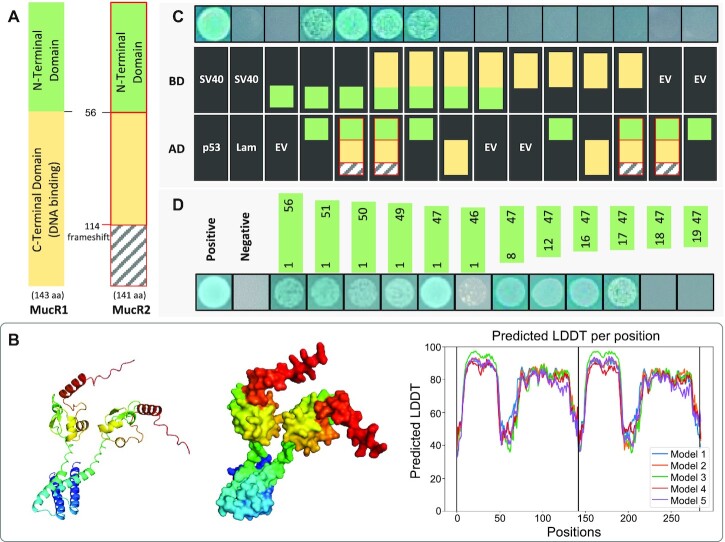
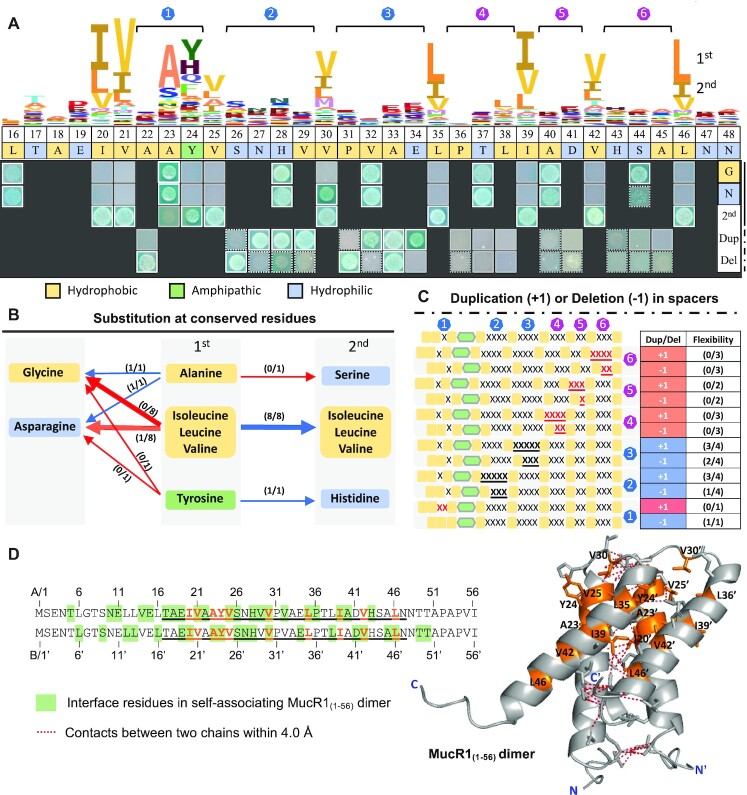
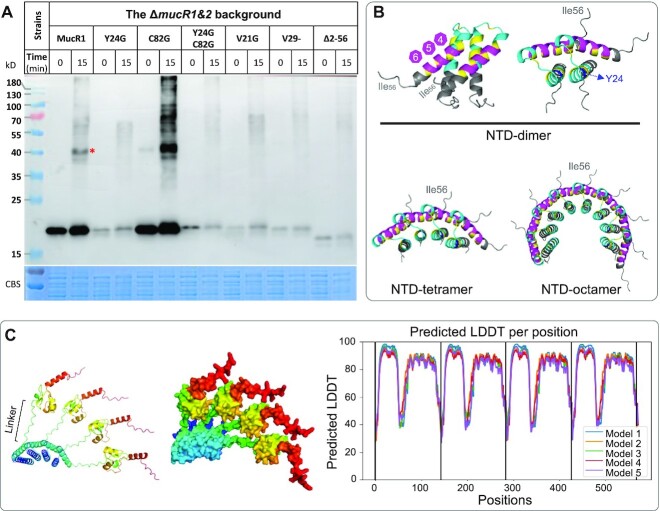
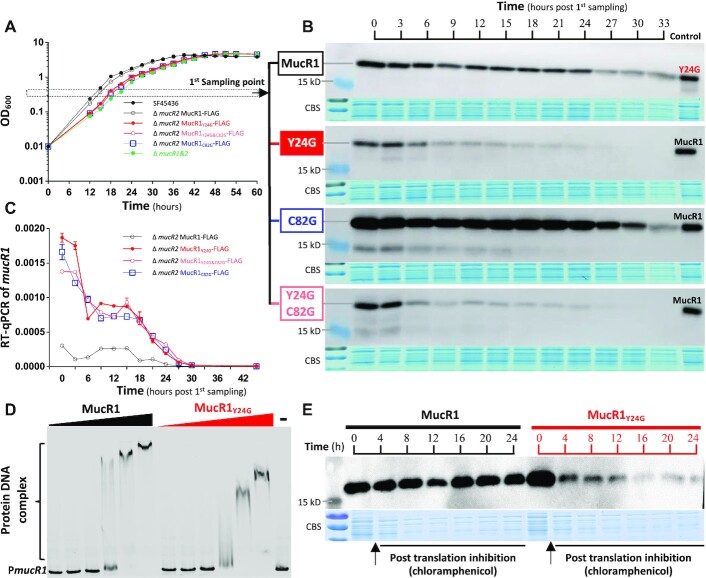
References
-
- López-García P., Moreira D.. Cultured Asgard archaea shed light on eukaryogenesis. Cell. 2020; 181:232–235. - PubMed
-
- Lane N. How energy flow shapes cell evolution. Curr. Biol. 2020; 30:R471–R476. - PubMed
-
- Brunk C.F., Martin W.F.. Archaeal histone contributions to the origin of eukaryotes. Trends Microbiol. 2019; 27:703–714. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
