

Oligodendrocyte differentiation alters tRNA modifications and codon optimality-mediated mRNA decay
- PMID: 36008413
- PMCID: PMC9411196
- DOI: 10.1038/s41467-022-32766-3
Oligodendrocyte differentiation alters tRNA modifications and codon optimality-mediated mRNA decay
Abstract
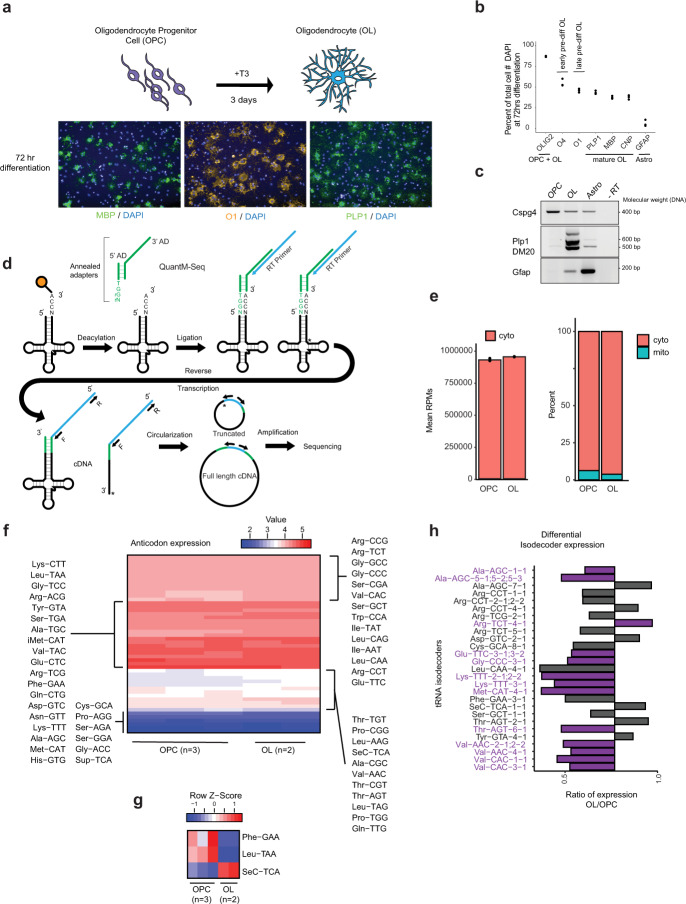
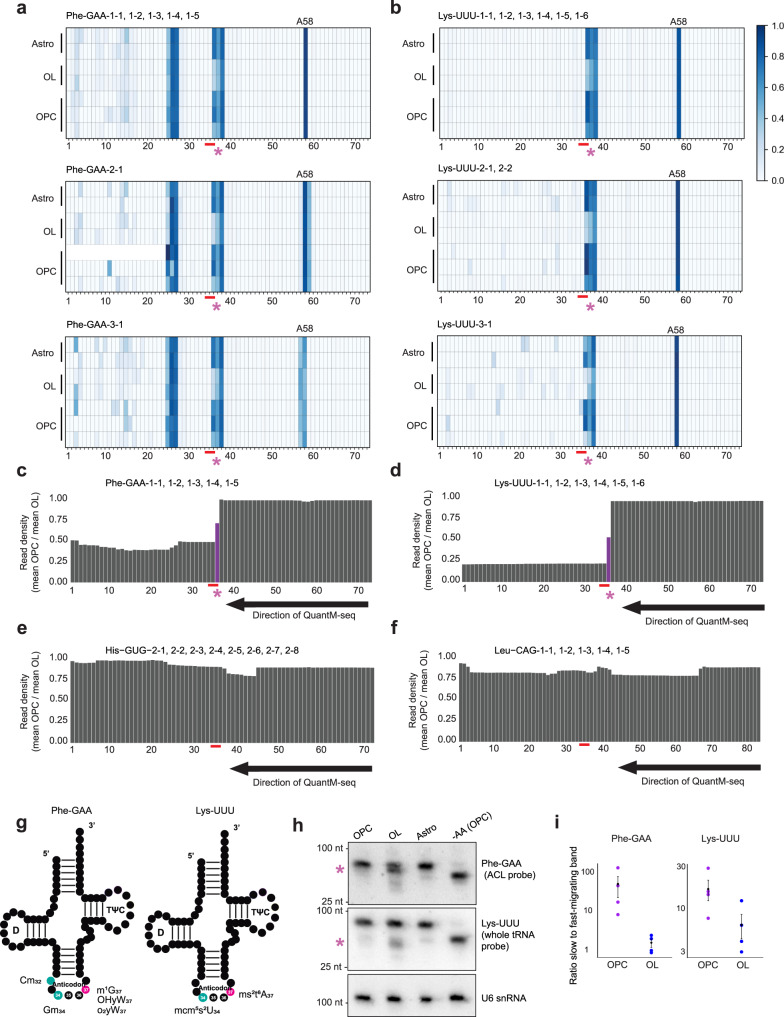
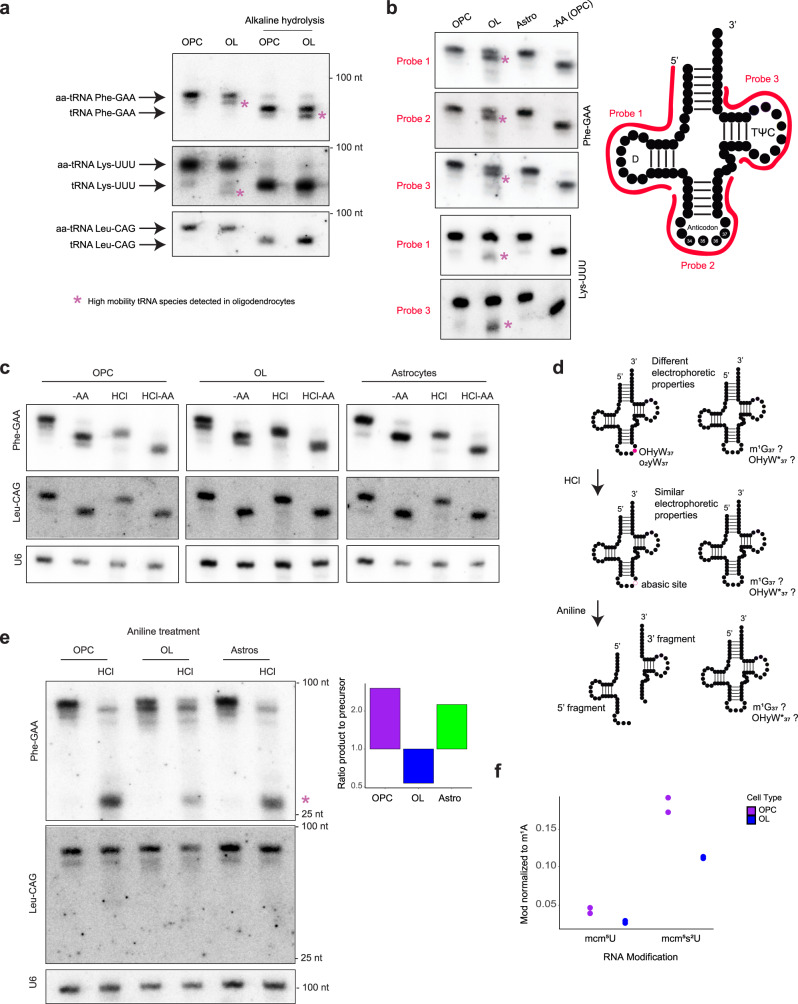
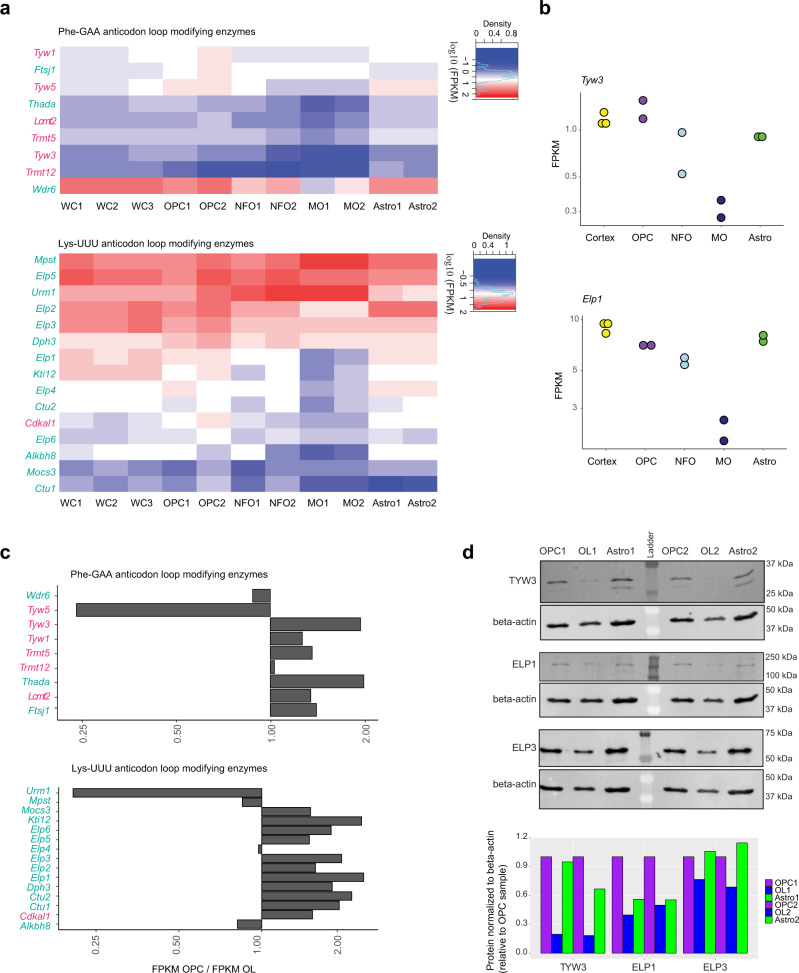
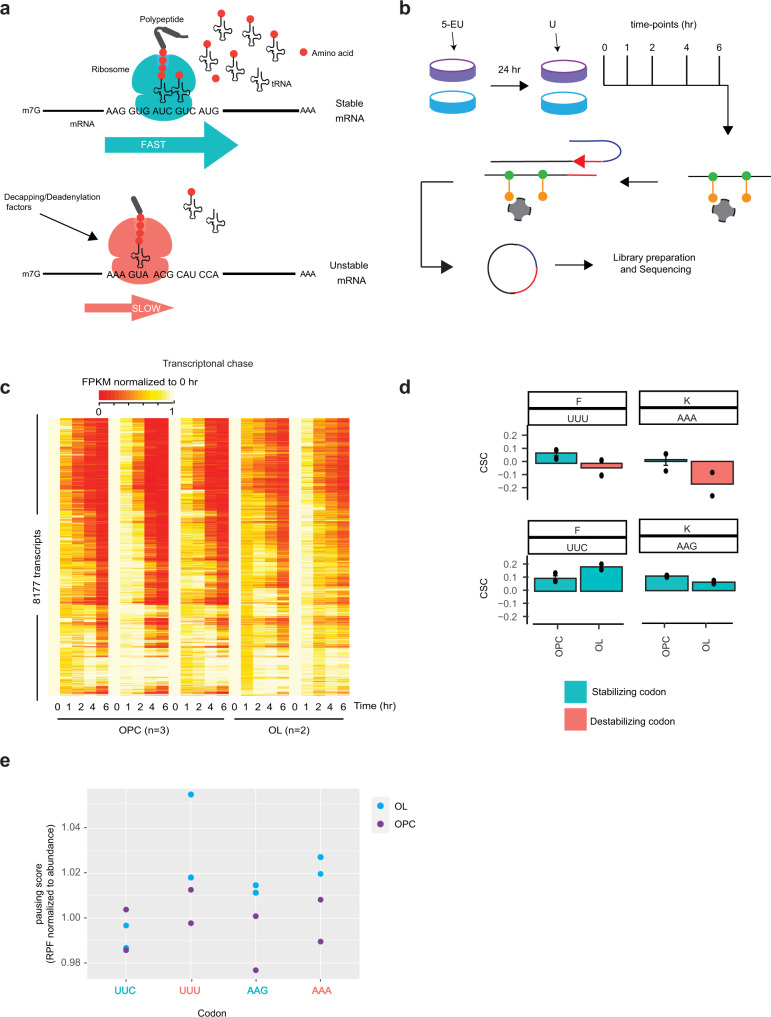
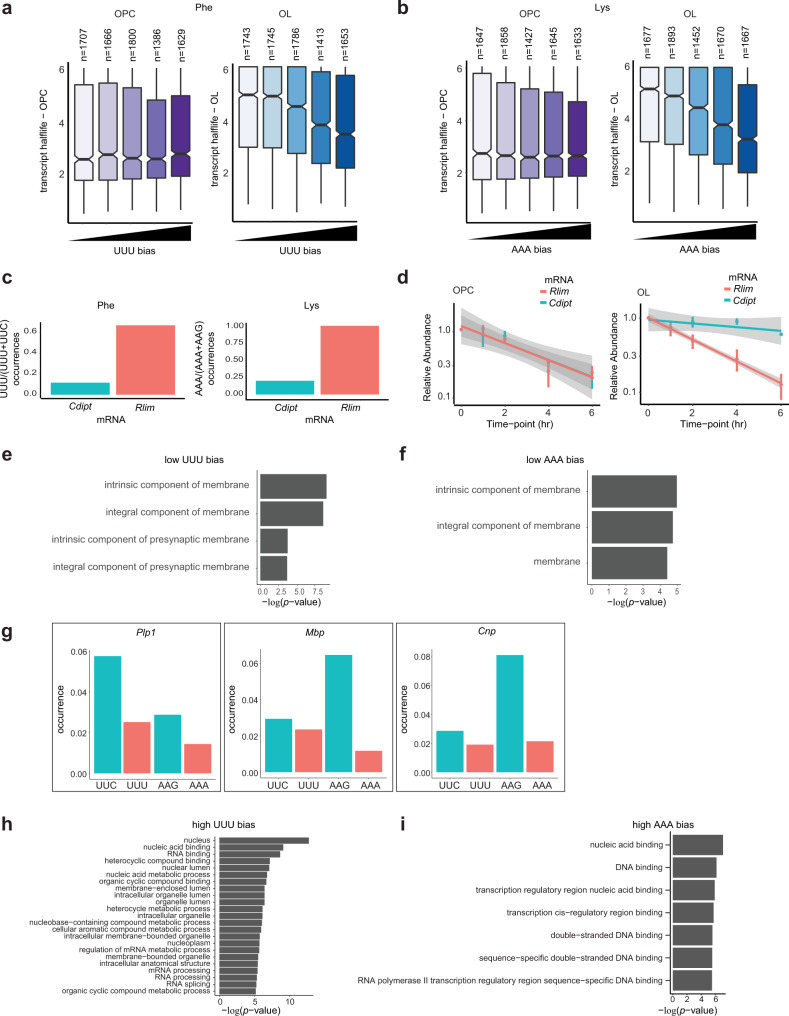
Oligodendrocytes are specialized cells that confer neuronal myelination in the central nervous system. Leukodystrophies associated with oligodendrocyte deficits and hypomyelination are known to result when a number of tRNA metabolism genes are mutated. Thus, for unknown reasons, oligodendrocytes may be hypersensitive to perturbations in tRNA biology. In this study, we survey the tRNA transcriptome in the murine oligodendrocyte cell lineage and find that specific tRNAs are hypomodified in oligodendrocytes within or near the anticodon compared to oligodendrocyte progenitor cells (OPCs). This hypomodified state may be the result of differential expression of key modification enzymes during oligodendrocyte differentiation. Moreover, we observe a concomitant relationship between tRNA hypomodification and tRNA decoding potential; observing oligodendrocyte specific alterations in codon optimality-mediated mRNA decay and ribosome transit. Our results reveal that oligodendrocytes naturally maintain a delicate, hypersensitized tRNA/mRNA axis. We suggest this axis is a potential mediator of pathology in leukodystrophies and white matter disease when further insult to tRNA metabolism is introduced.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Disruption of evolutionarily correlated tRNA elements impairs accurate decoding.Proc Natl Acad Sci U S A. 2020 Jul 14;117(28):16333-16338. doi: 10.1073/pnas.2004170117. Epub 2020 Jun 29. Proc Natl Acad Sci U S A. 2020. PMID: 32601241 Free PMC article.
-
Loss of Tuberous Sclerosis Complex1 in Adult Oligodendrocyte Progenitor Cells Enhances Axon Remyelination and Increases Myelin Thickness after a Focal Demyelination.J Neurosci. 2017 Aug 2;37(31):7534-7546. doi: 10.1523/JNEUROSCI.3454-16.2017. Epub 2017 Jul 10. J Neurosci. 2017. PMID: 28694334 Free PMC article.
-
Transfer RNA Levels Are Tuned to Support Differentiation During Drosophila Neurogenesis.Genes (Basel). 2024 Dec 15;15(12):1602. doi: 10.3390/genes15121602. Genes (Basel). 2024. PMID: 39766869 Free PMC article.
-
The Importance of Being Modified: The Role of RNA Modifications in Translational Fidelity.Enzymes. 2017;41:1-50. doi: 10.1016/bs.enz.2017.03.005. Epub 2017 Apr 22. Enzymes. 2017. PMID: 28601219 Free PMC article. Review.
-
The ribosome's response to codon-anticodon mismatches.Biochimie. 2006 Aug;88(8):1001-11. doi: 10.1016/j.biochi.2006.04.013. Epub 2006 May 12. Biochimie. 2006. PMID: 16716484 Review.
Cited by
-
Single-cell RNA-based phenotyping reveals a pivotal role of thyroid hormone receptor alpha for hypothalamic development.Development. 2023 Feb 1;150(3):dev201228. doi: 10.1242/dev.201228. Epub 2023 Jan 30. Development. 2023. PMID: 36715020 Free PMC article.
-
Orb2 enables rare-codon-enriched mRNA expression during Drosophila neuron differentiation.Nat Commun. 2024 Jun 20;15(1):5270. doi: 10.1038/s41467-024-48344-8. Nat Commun. 2024. PMID: 38902233 Free PMC article.
-
Human organoid model of pontocerebellar hypoplasia 2a recapitulates brain region-specific size differences.Dis Model Mech. 2024 Jul 1;17(7):dmm050740. doi: 10.1242/dmm.050740. Epub 2024 Jul 22. Dis Model Mech. 2024. PMID: 39034883 Free PMC article.
-
Transfer RNA levels are tuned to support differentiation during Drosophila neurogenesis.bioRxiv [Preprint]. 2024 Sep 6:2024.09.06.611608. doi: 10.1101/2024.09.06.611608. bioRxiv. 2024. Update in: Genes (Basel). 2024 Dec 15;15(12):1602. doi: 10.3390/genes15121602. PMID: 39282315 Free PMC article. Updated. Preprint.
-
Combining Nanopore direct RNA sequencing with genetics and mass spectrometry for analysis of T-loop base modifications across 42 yeast tRNA isoacceptors.Nucleic Acids Res. 2024 Oct 28;52(19):12074-12092. doi: 10.1093/nar/gkae796. Nucleic Acids Res. 2024. PMID: 39340295 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
