

Whole-Genome Identification of APX and CAT Gene Families in Cultivated and Wild Soybeans and Their Regulatory Function in Plant Development and Stress Response
- PMID: 36009347
- PMCID: PMC9404807
- DOI: 10.3390/antiox11081626
Whole-Genome Identification of APX and CAT Gene Families in Cultivated and Wild Soybeans and Their Regulatory Function in Plant Development and Stress Response
Erratum in
-
Correction: Aleem et al. Whole-Genome Identification of APX and CAT Gene Families in Cultivated and Wild Soybeans and Their Regulatory Function in Plant Development and Stress Response. Antioxidants 2022, 11, 1626.Antioxidants (Basel). 2025 Feb 18;14(2):229. doi: 10.3390/antiox14020229. Antioxidants (Basel). 2025. PMID: 40002429 Free PMC article.
Abstract
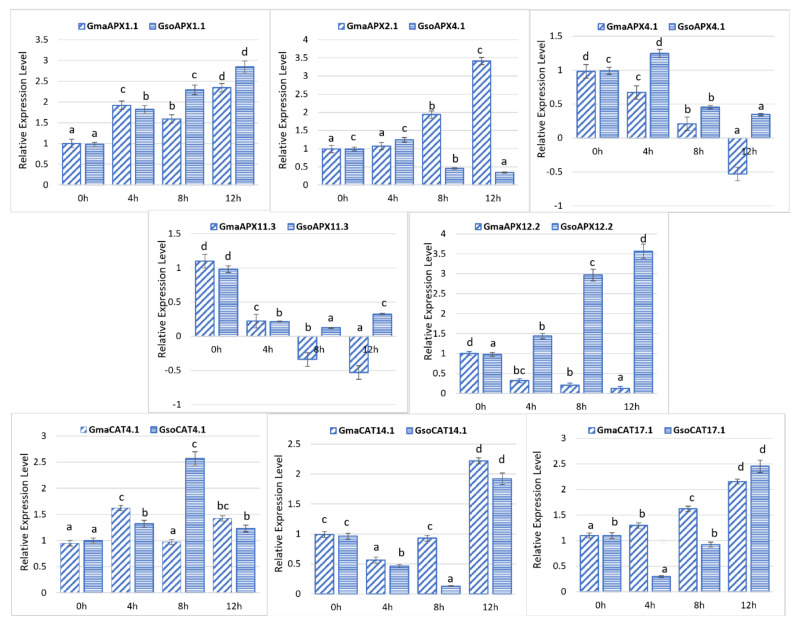
Plants coevolved with their antioxidant defense systems, which detoxify and adjust levels of reactive oxygen species (ROS) under multiple plant stresses. We performed whole-genome identification of ascorbate peroxidase (APX) and catalase (CAT) families in cultivated and wild soybeans. In cultivated and wild soybean genomes, we identified 11 and 10 APX genes, respectively, whereas the numbers of identified CAT genes were four in each species. Comparative phylogenetic analysis revealed more homology among cultivated and wild soybeans relative to other legumes. Exon/intron structure, motif and synteny blocks are conserved in cultivated and wild species. According to the Ka/Ks value, purifying selection is a major force for evolution of these gene families in wild soybean; however, the APX gene family was evolved by both positive and purifying selection in cultivated soybean. Segmental duplication was a major factor involved in the expansion of APX and CAT genes. Expression patterns revealed that APX and CAT genes are differentially expressed across fourteen different soybean tissues under water deficit (WD), heat stress (HS) and combined drought plus heat stress (WD + HS). Altogether, the current study provides broad insights into these gene families in soybeans. Our results indicate that APX and CAT gene families modulate multiple stress response in soybeans.
Keywords: G. max; G. soja; ascorbate peroxidase; catalyze; legumes.
Conflict of interest statement
The authors declare no conflict of interest.
Figures







References
-
- Ahmad S., Tang L., Shahzad R., Mawia A.M., Rao G.S., Jamil S., Wei C., Sheng Z., Shao G., Wei X. CRISPR-based crop improvements: A way forward to achieve zero hunger. J. Agric. Food Chem. 2021;69:8307–8323. - PubMed
-
- Shahzad R., Shakra Jamil S.A., Nisar A., Amina Z., Saleem S., Iqbal M.Z., Atif R.M., Wang X. Harnessing the potential of plant transcription factors in developing climate resilient crops to improve global food security: Current and future perspectives. Saudi J. Biol. Sci. 2021;28:2323. - PMC - PubMed
-
- Jamil S., Shahzad R., Ahmad S., Ali Z., Shaheen S., Shahzadee H., Fatima N., Sharif I., Aleem M., Sultana R. Climate Change and Role of Genetics and Genomics in Climate-Resilient Sorghum. In: Fahad S., Sonmez O., Saud S., Wang D., Wu C., Adnan M., Turan V., editors. Developing Climate-Resilient Crops. CRC Press; Boca Raton, FL, USA: 2021. pp. 111–138.
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
