

Adipose Tissue-Derived Mesenchymal Stromal/Stem Cells, Obesity and the Tumor Microenvironment of Breast Cancer
- PMID: 36010901
- PMCID: PMC9405791
- DOI: 10.3390/cancers14163908
Adipose Tissue-Derived Mesenchymal Stromal/Stem Cells, Obesity and the Tumor Microenvironment of Breast Cancer
Abstract
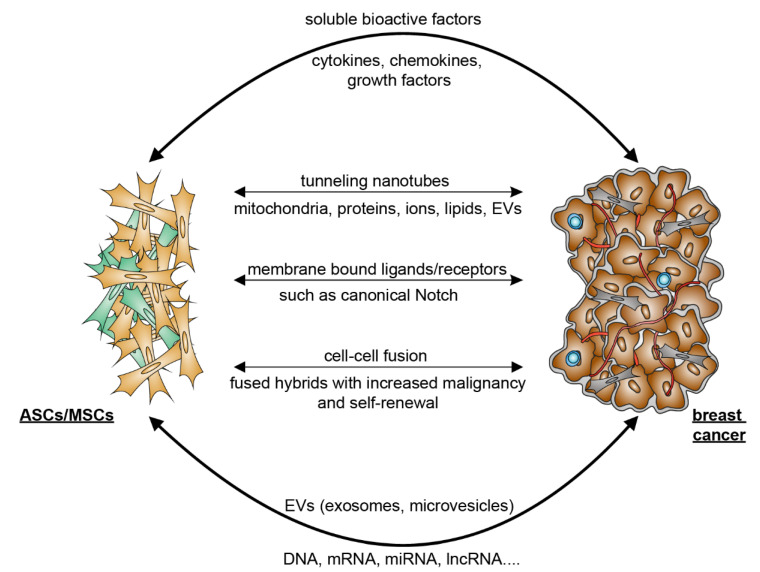
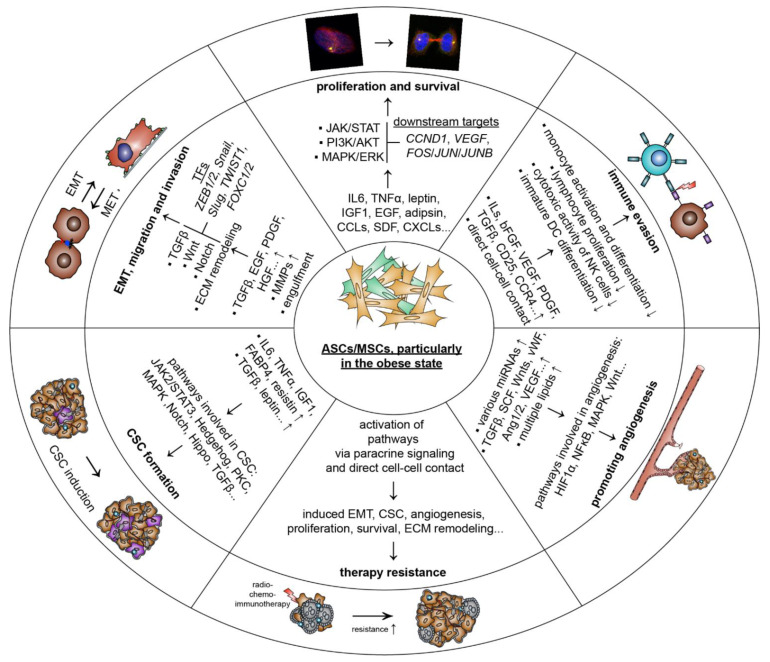
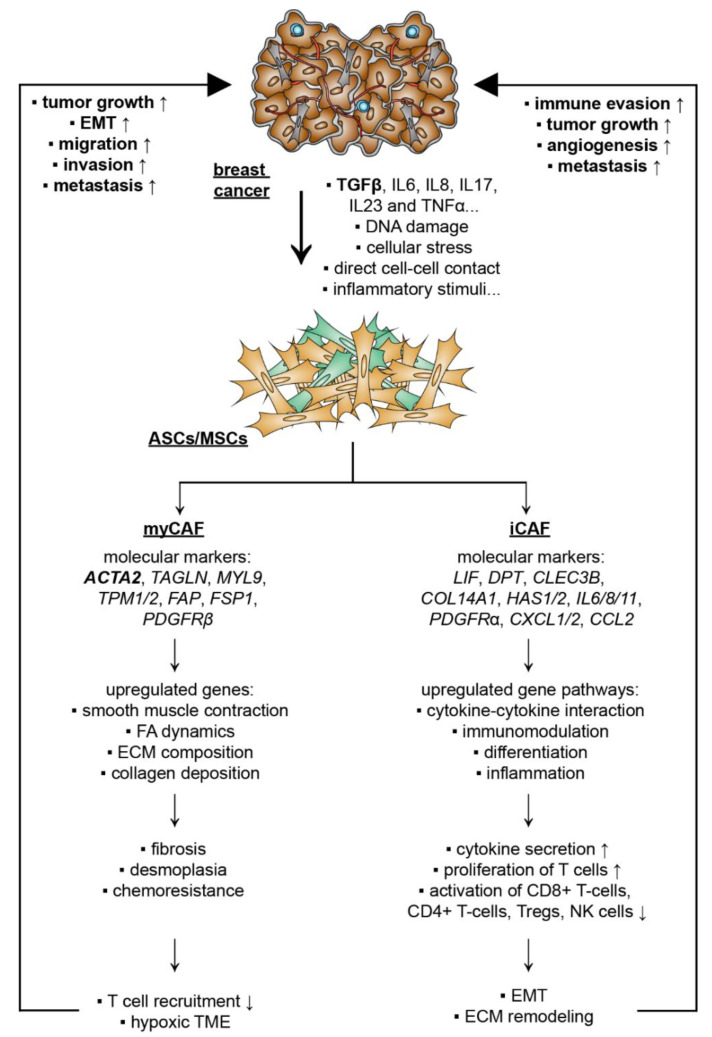
Breast cancer is the most frequently diagnosed cancer and a common cause of cancer-related death in women. It is well recognized that obesity is associated with an enhanced risk of more aggressive breast cancer as well as reduced patient survival. Adipose tissue is the major microenvironment of breast cancer. Obesity changes the composition, structure, and function of adipose tissue, which is associated with inflammation and metabolic dysfunction. Interestingly, adipose tissue is rich in ASCs/MSCs, and obesity alters the properties and functions of these cells. As a key component of the mammary stroma, ASCs play essential roles in the breast cancer microenvironment. The crosstalk between ASCs and breast cancer cells is multilateral and can occur both directly through cell-cell contact and indirectly via the secretome released by ASC/MSC, which is considered to be the main effector of their supportive, angiogenic, and immunomodulatory functions. In this narrative review, we aim to address the impact of obesity on ASCs/MSCs, summarize the current knowledge regarding the potential pathological roles of ASCs/MSCs in the development of breast cancer, discuss related molecular mechanisms, underline the possible clinical significance, and highlight related research perspectives. In particular, we underscore the roles of ASCs/MSCs in breast cancer cell progression, including proliferation and survival, angiogenesis, migration and invasion, the epithelial-mesenchymal transition, cancer stem cell development, immune evasion, therapy resistance, and the potential impact of breast cancer cells on ASCS/MSCs by educating them to become cancer-associated fibroblasts. We conclude that ASCs/MSCs, especially obese ASCs/MSCs, may be key players in the breast cancer microenvironment. Targeting these cells may provide a new path of effective breast cancer treatment.
Keywords: ASCs/MSCs; breast cancer; cancer-associated fibroblasts; cancer-associated stem cells; epithelial–mesenchymal transition; obesity; therapy resistance; tumor microenvironment.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Di Cesare M., Bentham J., Stevens G.A., Zhou B., Danaei G., Lu Y., Bixby H., Cowan M.J., Riley L.M., Hajifathalian K., et al. Trends in adult body-mass index in 200 countries from 1975 to 2014: A pooled analysis of 1698 population-based measurement studies with 19.2 million participants. Lancet. 2016;387:1377–1396. doi: 10.1016/s0140-6736(16)30054-x. - DOI - PMC - PubMed
-
- Bixby H., Bentham J., Zhou B., Di Cesare M., Paciorek C.J., Bennett J.E., Taddei C., Stevens G.A., Rodriguez-Martinez A., Carrillo-Larco R.M., et al. Rising rural body-mass index is the main driver of the global obesity epidemic in adults. Nature. 2019;569:260–264. doi: 10.1038/s41586-019-1171-x. - DOI - PMC - PubMed
-
- Wang L.M., Zhou B., Zhao Z.P., Yang L., Zhang M., Jiang Y., Li Y.C., Zhou M.G., Wang L.H., Huang Z.J., et al. Body-mass index and obesity in urban and rural China: Findings from consecutive nationally representative surveys during 2004–18. Lancet. 2021;398:53–63. doi: 10.1016/S0140-6736(21)00798-4. - DOI - PMC - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
