

Toxic and Phenotypic Effects of AAV_Cre Used to Transduce Mesencephalic Dopaminergic Neurons
- PMID: 36012727
- PMCID: PMC9408874
- DOI: 10.3390/ijms23169462
Toxic and Phenotypic Effects of AAV_Cre Used to Transduce Mesencephalic Dopaminergic Neurons
Abstract
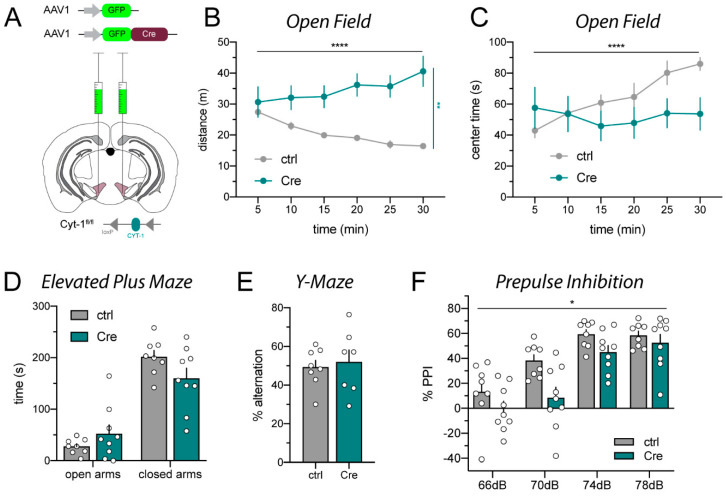
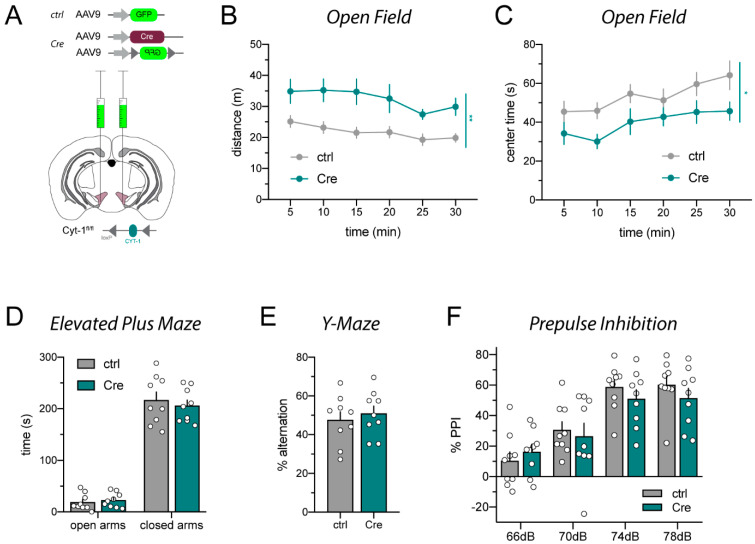
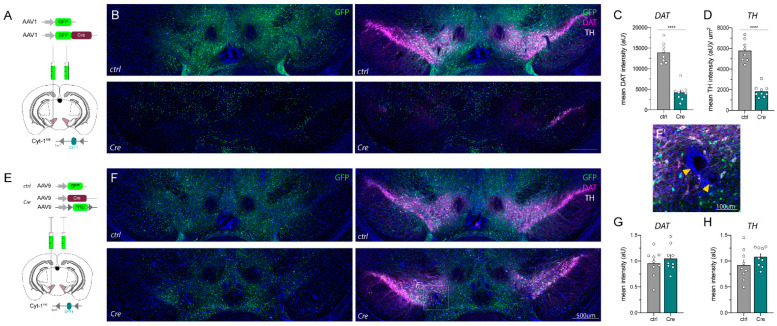
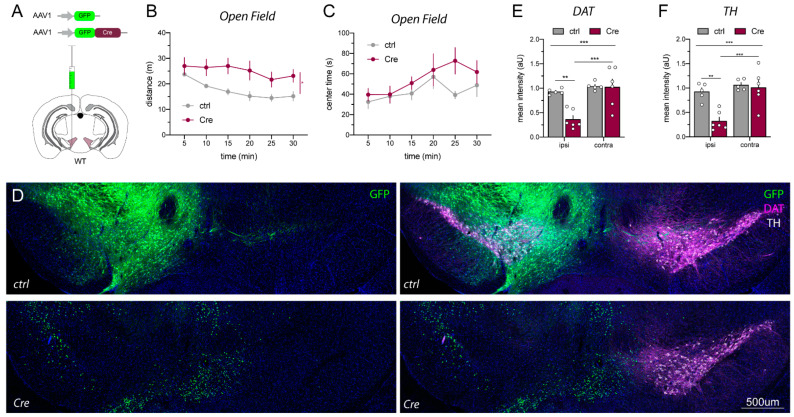
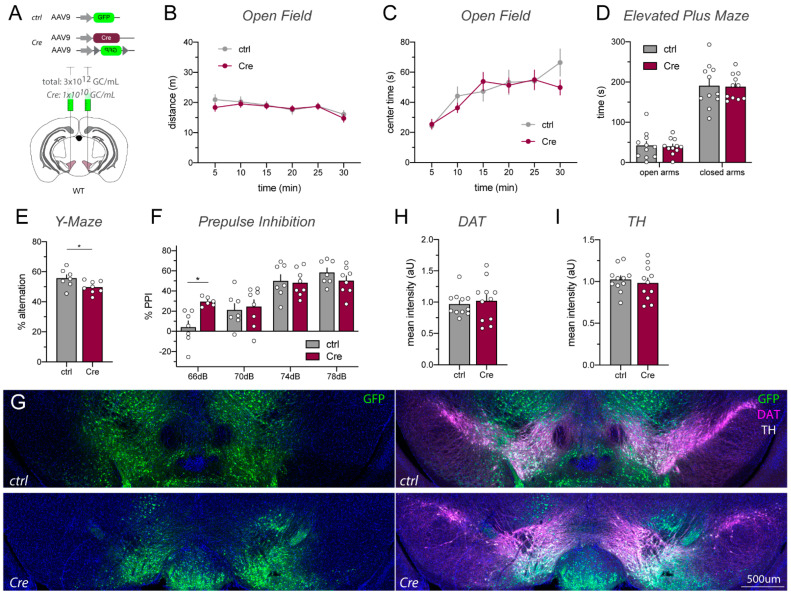
A popular approach to spatiotemporally target genes using the loxP/Cre recombination system is stereotaxic microinjection of adeno-associated virus (AAV) expressing Cre recombinase (AAV_Cre) in specific neuronal structures. Here, we report that AAV_Cre microinjection in the ventral tegmental area (VTA) of ErbB4 Cyt-1-floxed (ErbB4 Cyt-1fl/fl) mice at titers commonly used in the literature (~1012-1013 GC/mL) can have neurotoxic effects on dopaminergic neurons and elicit behavioral abnormalities. However, these effects of AAV_Cre microinjection are independent of ErbB4 Cyt-1 recombination because they are also observed in microinjected wild-type (WT) controls. Mice microinjected with AAV_Cre (1012-1013 GC/mL) exhibit reductions of tyrosine hydroxylase (TH) and dopamine transporter (DAT) expression, loss of dopaminergic neurons, and they behaviorally become hyperactive, fail to habituate in the open field and exhibit sensorimotor gating deficits compared to controls microinjected with AAV_GFP. Importantly, these AAV_Cre non-specific effects are: (1) independent of serotype, (2) occur with vectors expressing either Cre or Cre-GFP fusion protein and (3) preventable by reducing viral titers by 1000-fold (1010 GC/mL), which retains sufficient recombination activity to target floxed genes. Our studies emphasize the importance of including AAV_Cre-injected WT controls in experiments because recombination-independent effects on gene expression, neurotoxicity and behaviors could be erroneously attributed to consequences of gene ablation.
Keywords: Cre recombinase; adeno-associated virus; dopamine; stereotaxic injection; toxicity; tyrosine hydroxylase; ventral tegmental area.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
