

Dominantly acting KIF5B variants with pleiotropic cellular consequences cause variable clinical phenotypes
- PMID: 36018820
- PMCID: PMC9851748
- DOI: 10.1093/hmg/ddac213
Dominantly acting KIF5B variants with pleiotropic cellular consequences cause variable clinical phenotypes
Abstract
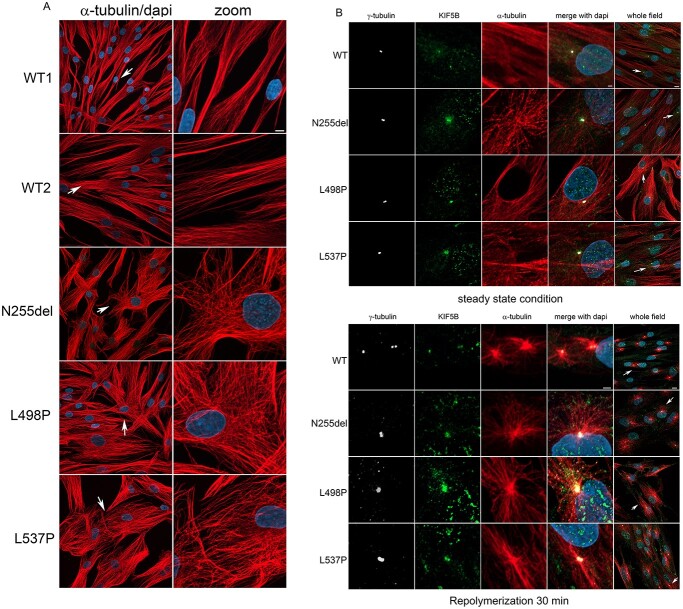
Kinesins are motor proteins involved in microtubule (MT)-mediated intracellular transport. They contribute to key cellular processes, including intracellular trafficking, organelle dynamics and cell division. Pathogenic variants in kinesin-encoding genes underlie several human diseases characterized by an extremely variable clinical phenotype, ranging from isolated neurodevelopmental/neurodegenerative disorders to syndromic phenotypes belonging to a family of conditions collectively termed as 'ciliopathies.' Among kinesins, kinesin-1 is the most abundant MT motor for transport of cargoes towards the plus end of MTs. Three kinesin-1 heavy chain isoforms exist in mammals. Different from KIF5A and KIF5C, which are specifically expressed in neurons and established to cause neurological diseases when mutated, KIF5B is an ubiquitous protein. Three de novo missense KIF5B variants were recently described in four subjects with a syndromic skeletal disorder characterized by kyphomelic dysplasia, hypotonia and DD/ID. Here, we report three dominantly acting KIF5B variants (p.Asn255del, p.Leu498Pro and p.Leu537Pro) resulting in a clinically wide phenotypic spectrum, ranging from dilated cardiomyopathy with adult-onset ophthalmoplegia and progressive skeletal myopathy to a neurodevelopmental condition characterized by severe hypotonia with or without seizures. In vitro and in vivo analyses provide evidence that the identified disease-associated KIF5B variants disrupt lysosomal, autophagosome and mitochondrial organization, and impact cilium biogenesis. All variants, and one of the previously reported missense changes, were shown to affect multiple developmental processes in zebrafish. These findings document pleiotropic consequences of aberrant KIF5B function on development and cell homeostasis, and expand the phenotypic spectrum resulting from altered kinesin-mediated processes.
© The Author(s) 2022. Published by Oxford University Press. All rights reserved. For Permissions, please email: journals.permissions@oup.com.
Figures








References
-
- Hirokawa, N., Noda, Y., Tanaka, Y. and Niwa, S. (2009) Kinesin superfamily motor proteins and intracellular transport. Nat. Rev. Mol. Cell Biol., 10, 682–696. - PubMed
-
- Hirokawa, N. and Noda, Y. (2008) Intracellular transport and kinesin superfamily proteins, KIFs: structure, function, and dynamics. Physiol., 88, 1089–1118. - PubMed
-
- Verhey, K.J., Kaul, N. and Soppina, V. (2011) Kinesin assembly and movement in cells. Annu. Rev. Biophys., 40, 267–288. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
