

Exploring the mechanistic link between SF3B1 mutation and ring sideroblast formation in myelodysplastic syndrome
- PMID: 36028755
- PMCID: PMC9418223
- DOI: 10.1038/s41598-022-18921-2
Exploring the mechanistic link between SF3B1 mutation and ring sideroblast formation in myelodysplastic syndrome
Abstract
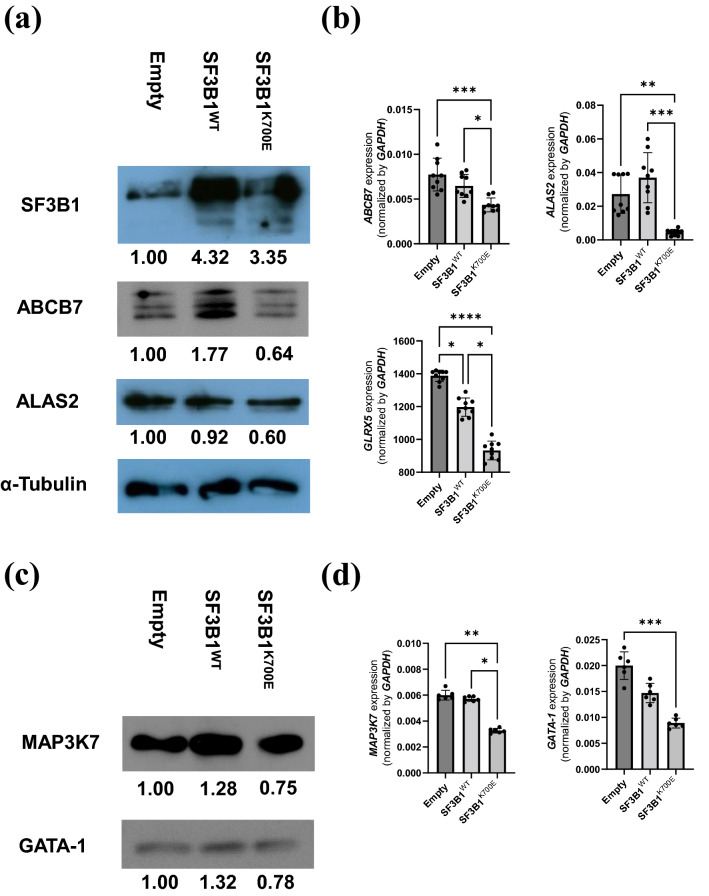
Acquired sideroblastic anemia, characterized by bone marrow ring sideroblasts (RS), is predominantly associated with myelodysplastic syndrome (MDS). Although somatic mutations in splicing factor 3b subunit 1 (SF3B1), which is involved in the RNA splicing machinery, are frequently found in MDS-RS, the detailed mechanism contributing to RS formation is unknown. To explore the mechanism, we established human umbilical cord blood-derived erythroid progenitor-2 (HUDEP-2) cells stably expressing SF3B1K700E. SF3B1K700E expressing cells showed higher proportion of RS than the control cells along with erythroid differentiation, indicating the direct contribution of mutant SF3B1 expression in erythroblasts to RS formation. In SF3B1K700E expressing cells, ABCB7 and ALAS2, known causative genes for congenital sideroblastic anemia, were downregulated. Additionally, mis-splicing of ABCB7 was observed in SF3B1K700E expressing cells. ABCB7-knockdown HUDEP-2 cells revealed an increased frequency of RS formation along with erythroid differentiation, demonstrating the direct molecular link between ABCB7 defects and RS formation. ALAS2 protein levels were obviously decreased in ABCB7-knockdown cells, indicating decreased ALAS2 translation owing to impaired Fe-S cluster export by ABCB7 defects. Finally, RNA-seq analysis of MDS clinical samples demonstrated decreased expression of ABCB7 by the SF3B1 mutation. Our findings contribute to the elucidation of the complex mechanisms of RS formation in MDS-RS.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures







References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
