

RIPK3 dampens mitochondrial bioenergetics and lipid droplet dynamics in metabolic liver disease
- PMID: 36029129
- PMCID: PMC10026966
- DOI: 10.1002/hep.32756
RIPK3 dampens mitochondrial bioenergetics and lipid droplet dynamics in metabolic liver disease
Abstract
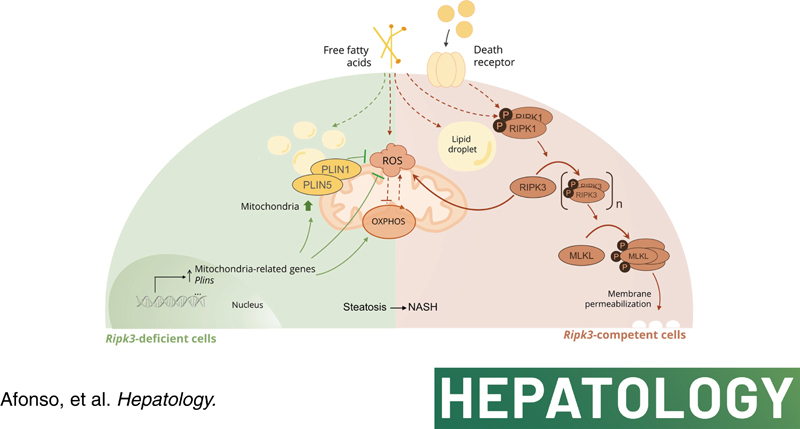
Background and aims: Receptor-interacting protein kinase 3 (RIPK3) mediates NAFLD progression, but its metabolic function is unclear. Here, we aimed to investigate the role of RIPK3 in modulating mitochondria function, coupled with lipid droplet (LD) architecture in NAFLD.
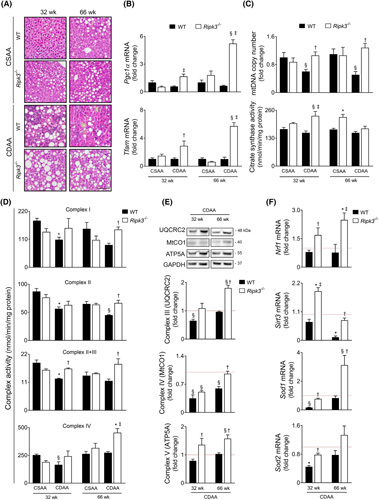
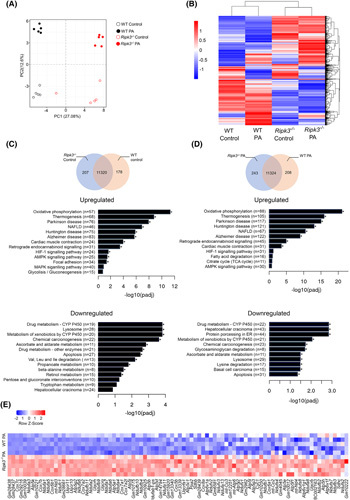
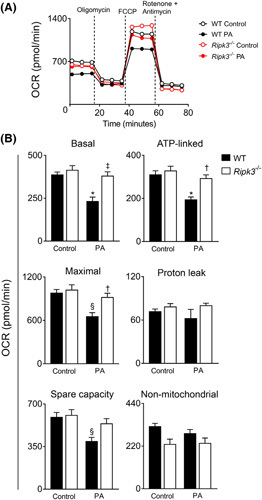
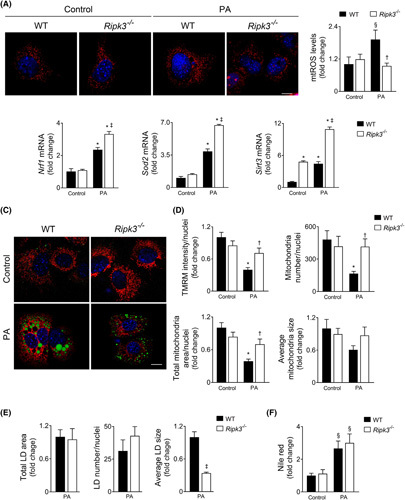
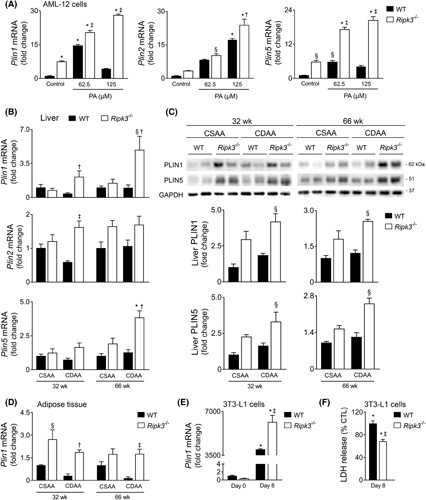
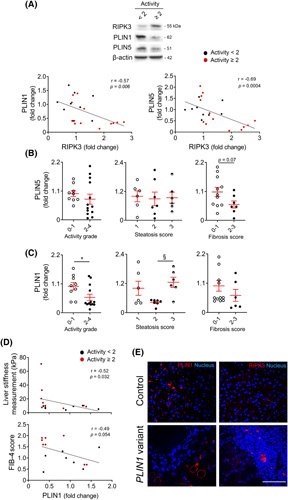
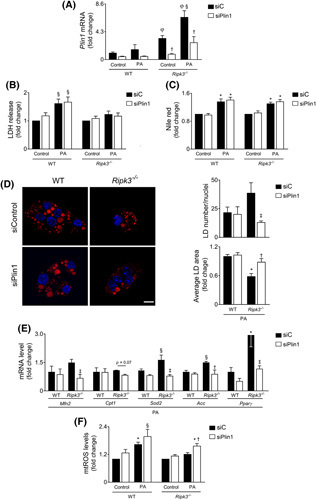
Approach and results: Functional studies evaluating mitochondria and LD biology were performed in wild-type (WT) and Ripk3-/- mice fed a choline-deficient, amino acid-defined (CDAA) diet for 32 and 66 weeks and in CRISPR-Cas9 Ripk3 -null fat-loaded immortalized hepatocytes. The association between hepatic perilipin (PLIN) 1 and 5, RIPK3, and disease severity was also addressed in a cohort of patients with NAFLD and in PLIN1 -associated familial partial lipodystrophy. Ripk3 deficiency rescued impairment in mitochondrial biogenesis, bioenergetics, and function in CDAA diet-fed mice and fat-loaded hepatocytes. Ripk3 deficiency was accompanied by a strong upregulation of antioxidant systems, leading to diminished oxidative stress upon fat loading both in vivo and in vitro. Strikingly, Ripk3-/- hepatocytes displayed smaller size LD in higher numbers than WT cells after incubation with free fatty acids. Ripk3 deficiency upregulated adipocyte and hepatic levels of LD-associated proteins PLIN1 and PLIN5. PLIN1 upregulation controlled LD structure and diminished mitochondrial stress upon free fatty acid overload in Ripk3-/- hepatocytes and was associated with diminished human NAFLD severity. Conversely, a pathogenic PLIN1 frameshift variant was associated with NAFLD and fibrosis, as well as with increased hepatic RIPK3 levels in familial partial lipodystrophy.
Conclusions: Ripk3 deficiency restores mitochondria bioenergetics and impacts LD dynamics. RIPK3 inhibition is promising in ameliorating NAFLD.
Copyright © 2023 The Author(s). Published by Wolters Kluwer Health, Inc.
Conflict of interest statement
The authors have no conflicts to report.
Figures
References
-
- Tilg H, Moschen AR. Evolution of inflammation in nonalcoholic fatty liver disease: the multiple parallel hits hypothesis. Hepatology. 2010;52:1836–46. - PubMed
-
- Afonso MB, Rodrigues PM, Carvalho T, Caridade M, Borralho P, Cortez‐Pinto H, et al. Necroptosis is a key pathogenic event in human and experimental murine models of non‐alcoholic steatohepatitis. Clin Sci (Lond). 2015;129:721–39. - PubMed
-
- Majdi A, Aoudjehane L, Ratziu V, Islam T, Afonso MB, Conti F, et al. Inhibition of receptor‐interacting protein kinase 1 improves experimental non‐alcoholic fatty liver disease. J Hepatol. 2020;72:627–35. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
