

Biallelic pathogenic variants in COX11 are associated with an infantile-onset mitochondrial encephalopathy
- PMID: 36030551
- PMCID: PMC9771894
- DOI: 10.1002/humu.24453
Biallelic pathogenic variants in COX11 are associated with an infantile-onset mitochondrial encephalopathy
Abstract
Primary mitochondrial diseases are a group of genetically and clinically heterogeneous disorders resulting from oxidative phosphorylation (OXPHOS) defects. COX11 encodes a copper chaperone that participates in the assembly of complex IV and has not been previously linked to human disease. In a previous study, we identified that COX11 knockdown decreased cellular adenosine triphosphate (ATP) derived from respiration, and that ATP levels could be restored with coenzyme Q10 (CoQ10 ) supplementation. This finding is surprising since COX11 has no known role in CoQ10 biosynthesis. Here, we report a novel gene-disease association by identifying biallelic pathogenic variants in COX11 associated with infantile-onset mitochondrial encephalopathies in two unrelated families using trio genome and exome sequencing. Functional studies showed that mutant COX11 fibroblasts had decreased ATP levels which could be rescued by CoQ10 . These results not only suggest that COX11 variants cause defects in energy production but reveal a potential metabolic therapeutic strategy for patients with COX11 variants.
Keywords: COX11; OXPHOS; coenzyme Q; mitochondrial disorders.
© 2022 The Authors. Human Mutation published by Wiley Periodicals LLC.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
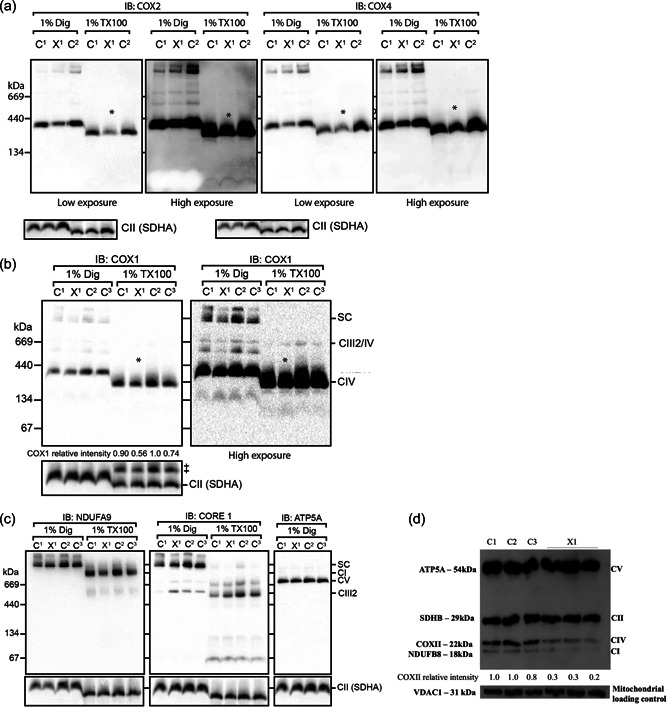
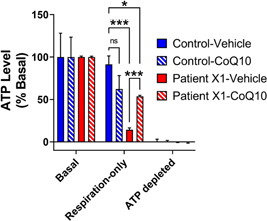
References
-
- DePristo, M. A. , Banks, E. , Poplin, R. , Garimella, K. V. , Maguire, J. R. , Hartl, C. , Philippakis, A. A. , del Angel, G. , Rivas, M. A. , Hanna, M. , McKenna, A. , Fennell, T. J. , Kernytsky, A. M. , Sivachenko, A. Y. , Cibulskis, K. , Gabriel, S. B. , Altshuler, D. , & Daly, M. J. (2011, May). A framework for variation discovery and genotyping using next‐generation DNA sequencing data. Nature Genetics, 43(5), 491–498. 10.1038/ng.806 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
