

Active-Site Engineering Switches Carbohydrate Regiospecificity in a Fungal Copper Radical Oxidase
- PMID: 36033369
- PMCID: PMC9397409
- DOI: 10.1021/acscatal.2c01956
Active-Site Engineering Switches Carbohydrate Regiospecificity in a Fungal Copper Radical Oxidase
Abstract
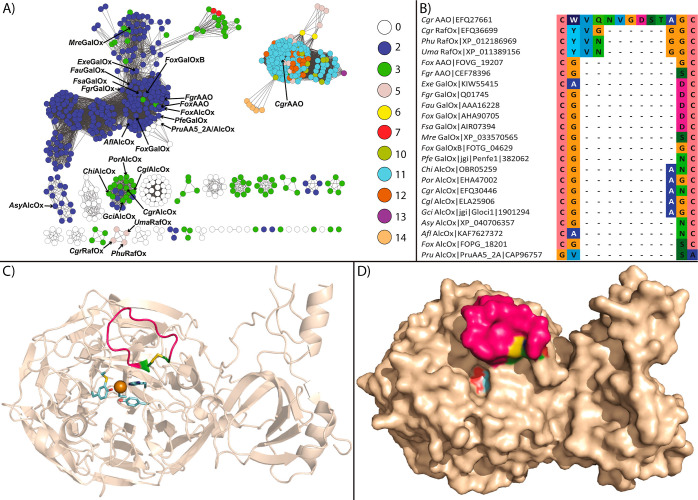
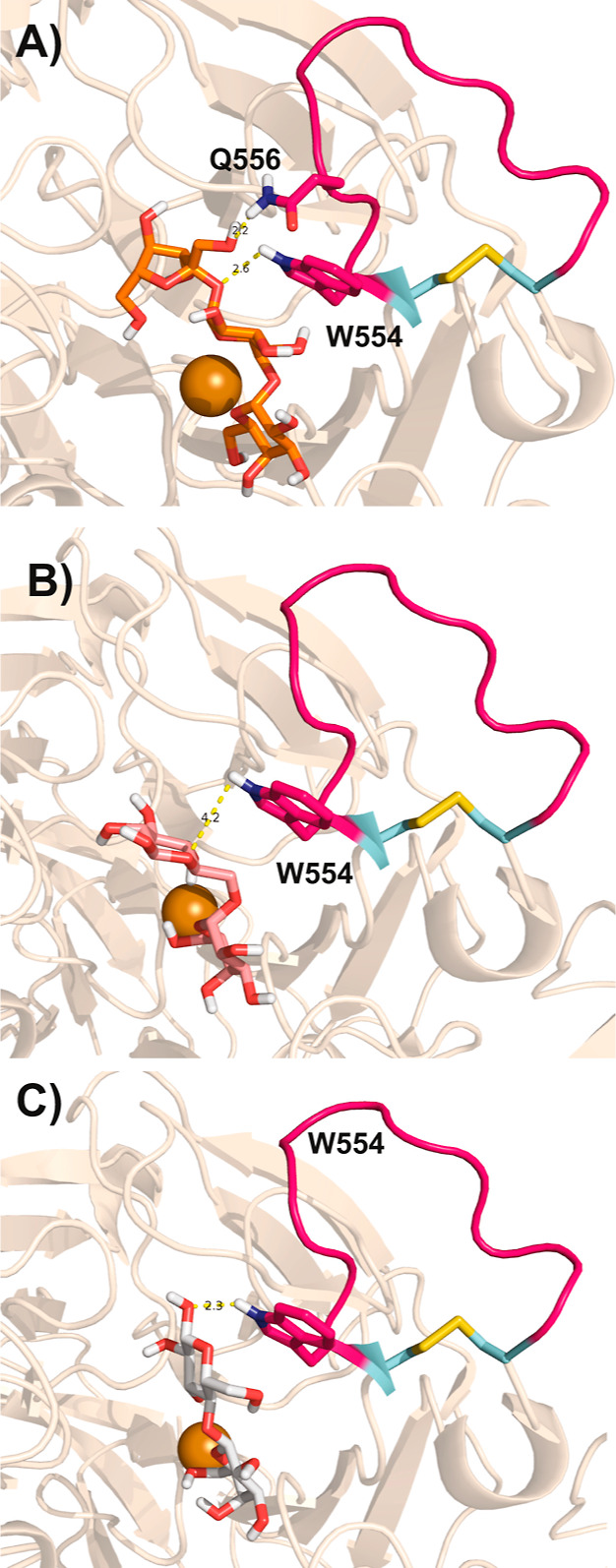
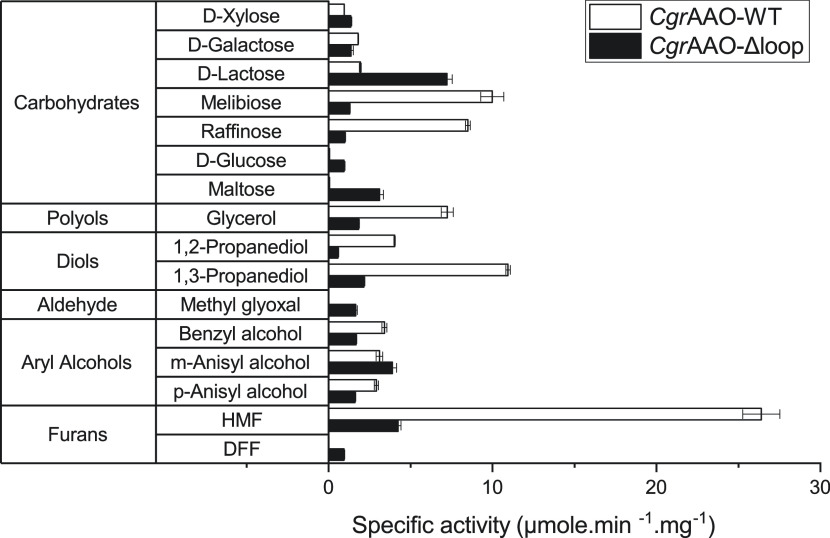
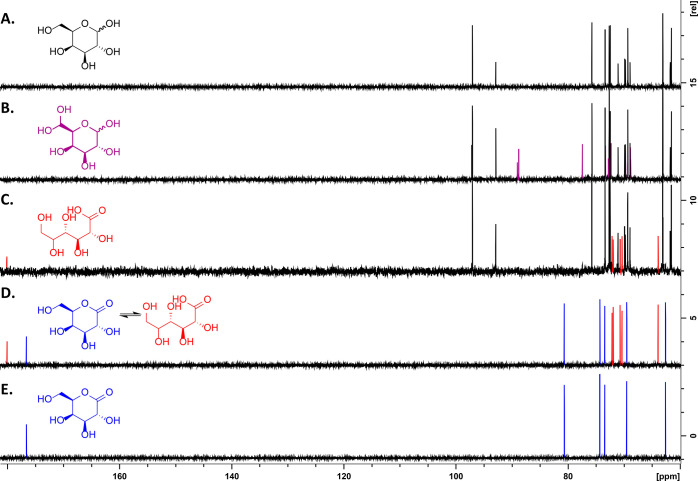
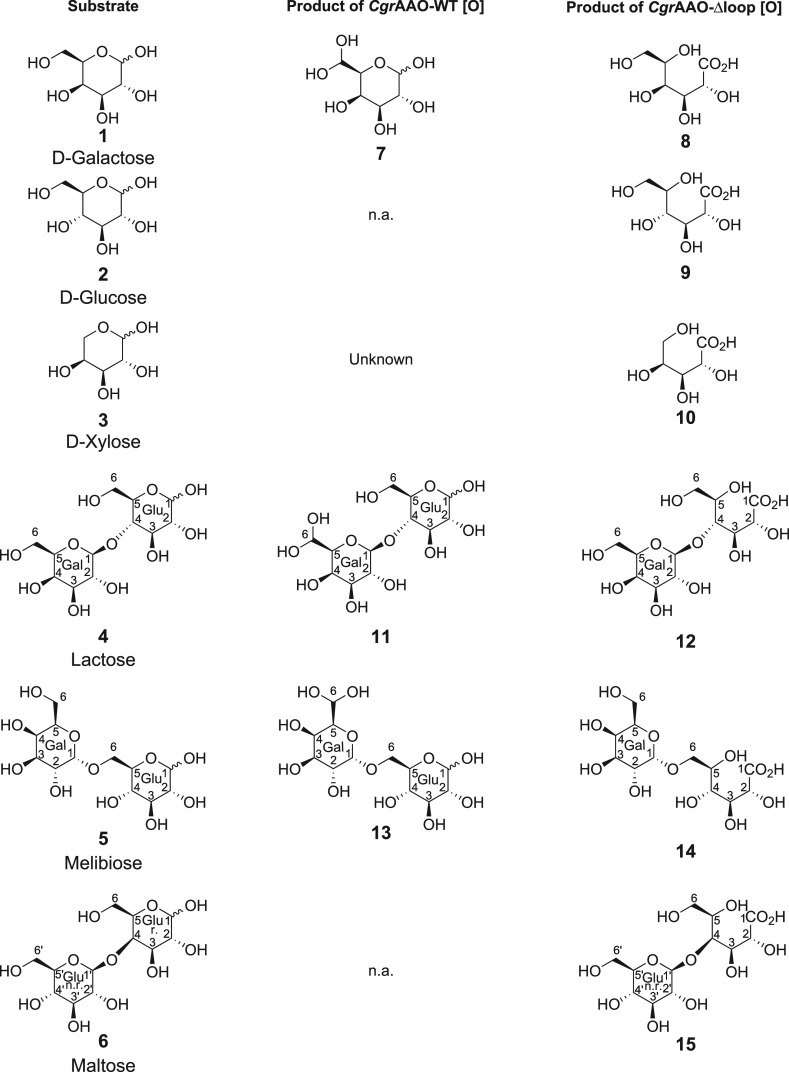
Copper radical oxidases (CROs) from Auxiliary Activity Family 5, Subfamily 2 (AA5_2), are organic cofactor-free biocatalysts for the selective oxidation of alcohols to the corresponding aldehydes. AA5_2 CROs comprise canonical galactose-6-oxidases as well as the more recently discovered general alcohol oxidases and aryl alcohol oxidases. Guided by primary and tertiary protein structural analyses, we targeted a distinct extended loop in the active site of a Colletotrichum graminicola aryl alcohol oxidase (CgrAAO) to explore its effect on catalysis in the broader context of AA5_2. Deletion of this loop, which is bracketed by a conserved disulfide bridge, significantly reduced the inherent activity of the enzyme toward extended galacto-oligosaccharides, as anticipated from molecular modeling. Unexpectedly, kinetic and product analysis on a range of monosaccharides and disaccharides revealed that an altered carbohydrate specificity in CgrAAO-Δloop was accompanied by a complete change in regiospecificity from C-6 to C-1 oxidation, thereby generating aldonic acids. C-1 regiospecificity is unprecedented in AA5 enzymes and is classically associated with flavin-dependent carbohydrate oxidases of Auxiliary Activity Family 3. Thus, this work further highlights the catalytic adaptability of the unique mononuclear copper radical active site and provides a basis for the design of improved biocatalysts for diverse potential applications.
© 2022 The Authors. Published by American Chemical Society.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
References
-
- Cooper J. A. D.; Smith W.; Bacila M.; Medina H. Galactose Oxidase from Polyporus circinatus. J. Biol. Chem. 1959, 234, 445–448. 10.1016/S0021-9258(18)70223-8. - DOI - PubMed
- Nobles M. K.; Madhosingh C. Dactyliumdendroides (Bull.) Fr. Misnamed as Polyporuscircinatus Fr. Biochem. Biophys. Res. Commun. 1963, 12, 146–147. 10.1016/0006-291X(63)90251-1. - DOI
LinkOut - more resources
Full Text Sources