

FXR Signaling-Mediated Bile Acid Metabolism Is Critical for Alleviation of Cholesterol Gallstones by Lactobacillus Strains
- PMID: 36036629
- PMCID: PMC9603329
- DOI: 10.1128/spectrum.00518-22
FXR Signaling-Mediated Bile Acid Metabolism Is Critical for Alleviation of Cholesterol Gallstones by Lactobacillus Strains
Abstract
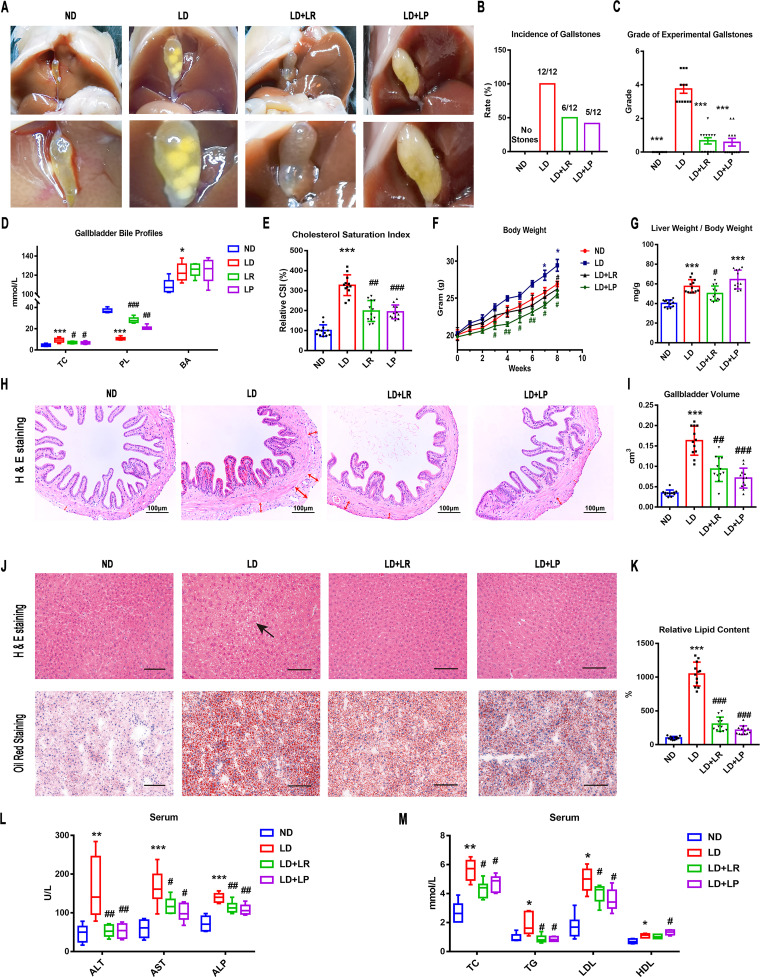
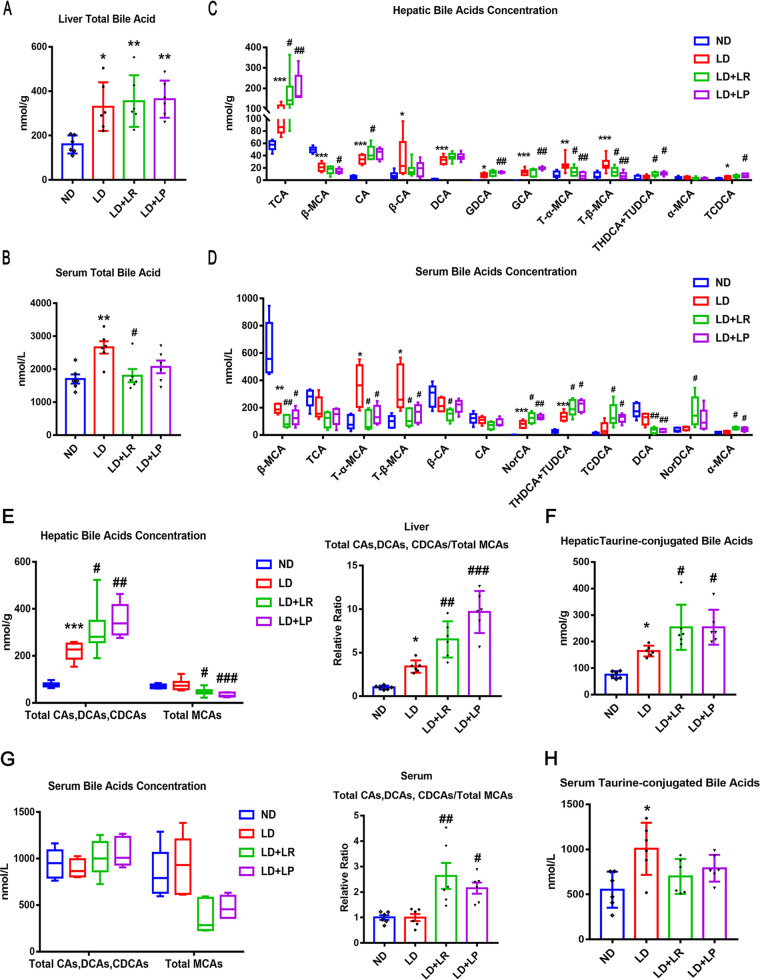
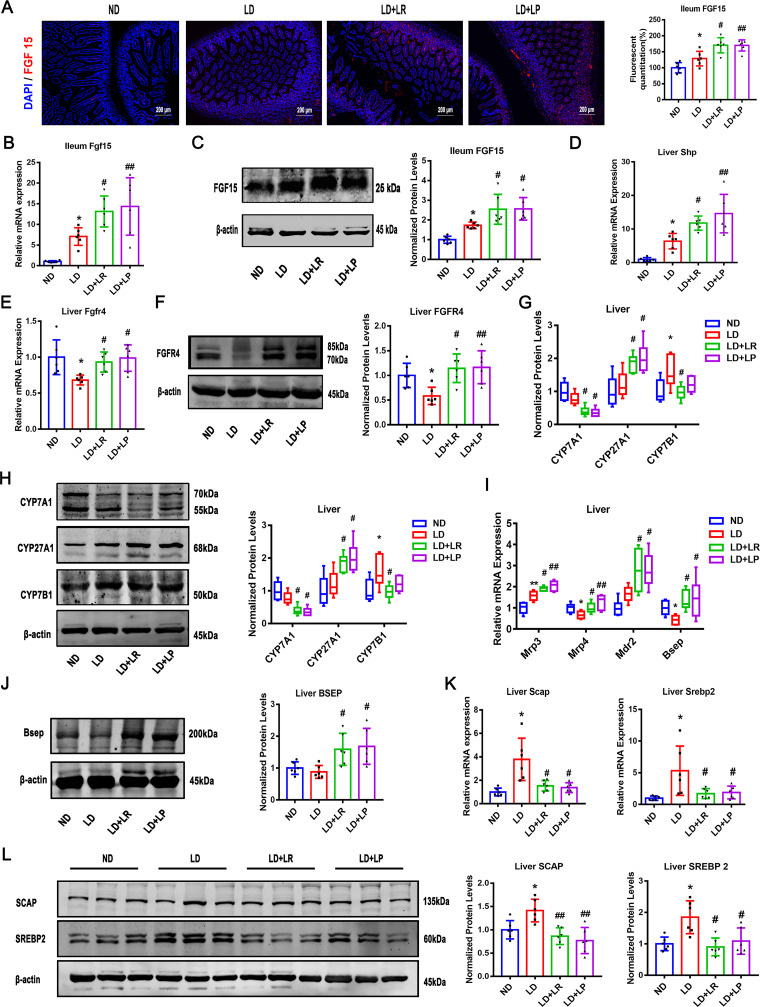
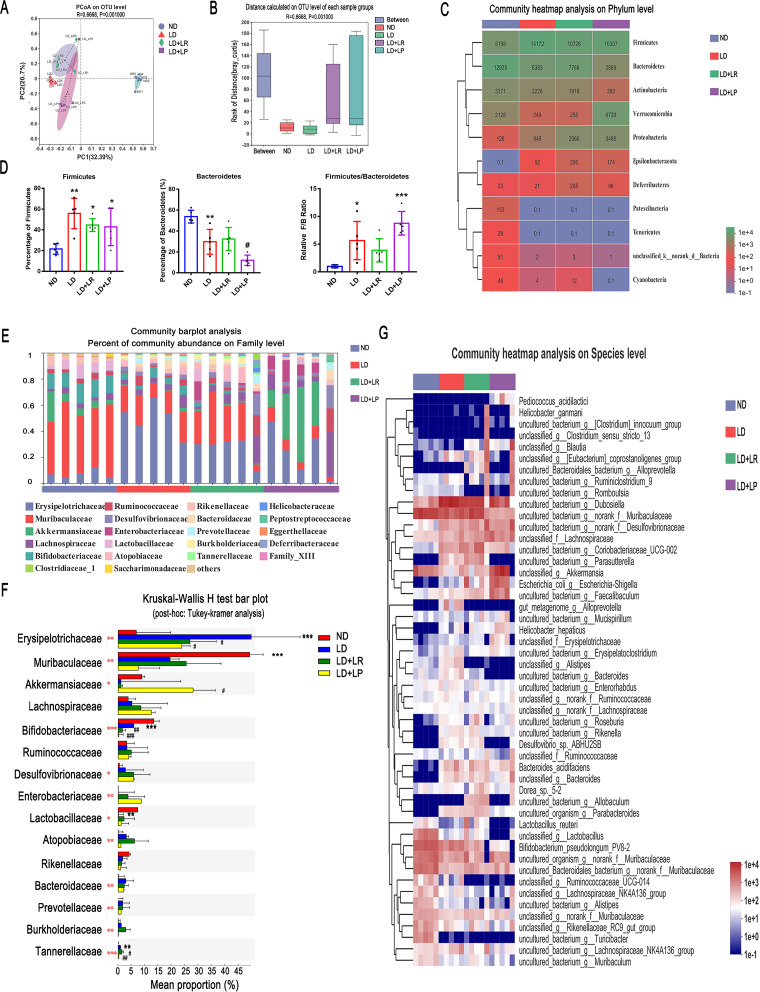
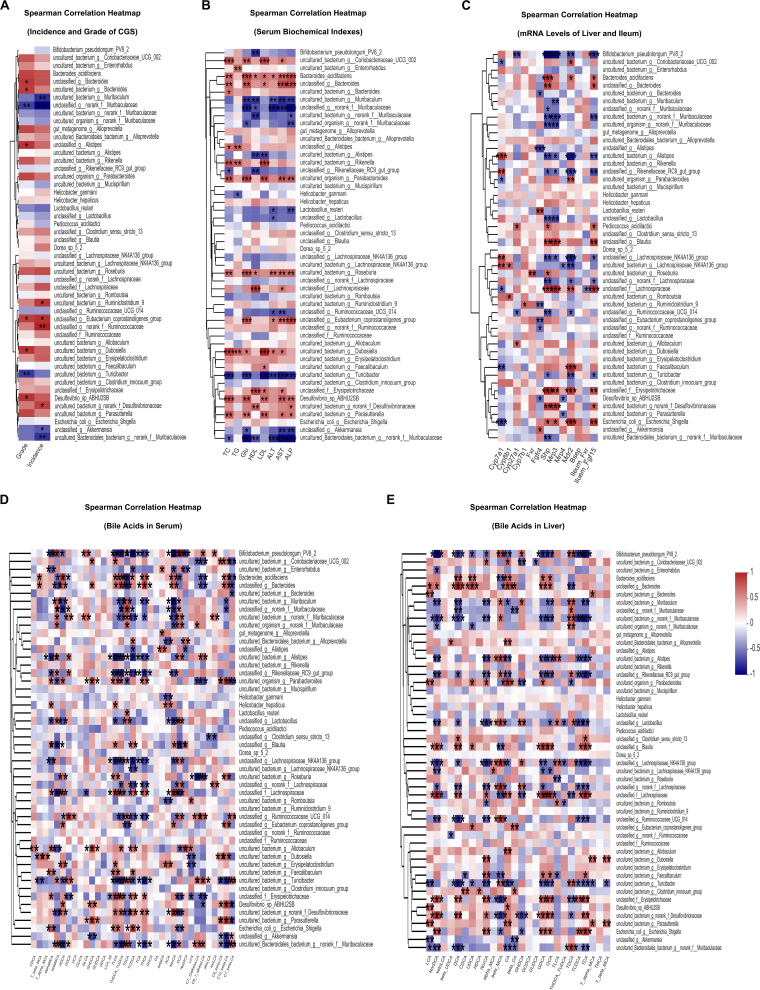
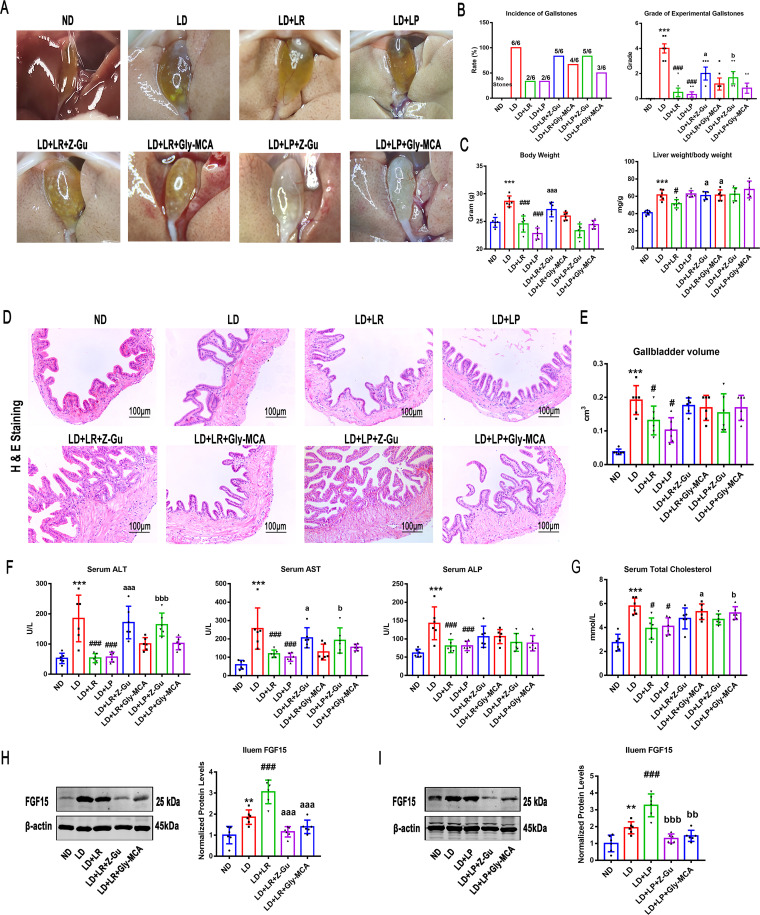
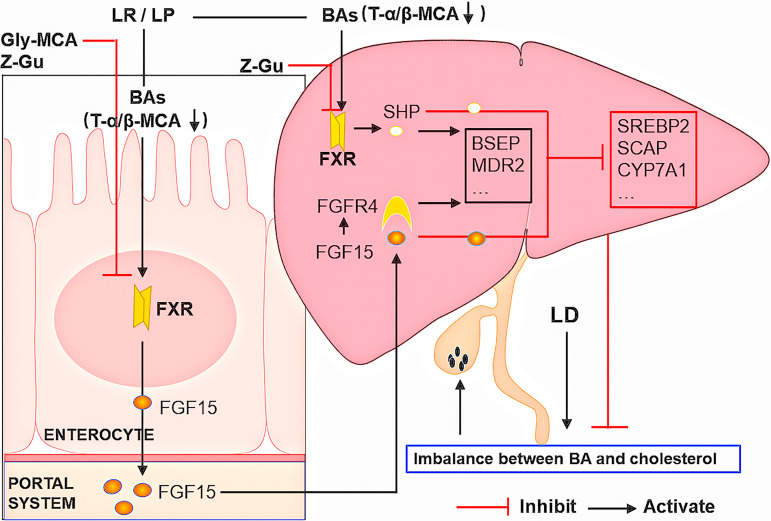
Cholesterol gallstone (CGS) disease is characterized by an imbalance in bile acid (BA) metabolism and is closely associated with gut microbiota disorders. However, the role and mechanism by which probiotics targeting the gut microbiota attenuate cholesterol gallstones are still unknown. In this study, Limosilactobacillus reuteri strain CGMCC 17942 and Lactiplantibacillus plantarum strain CGMCC 14407 were individually administered to lithogenic-diet (LD)-fed mice for 8 weeks. Both Lactobacillus strains significantly reduced LD-induced gallstones, hepatic steatosis, and hyperlipidemia. These strains modulated BA profiles in the serum and liver, which may be responsible for the activation of farnesoid X receptor (FXR). At the molecular level, L. reuteri and L. plantarum increased ileal fibroblast growth factor 15 (FGF15) and hepatic fibroblast growth factor receptor 4 (FGFR4) and small heterodimer partner (SHP). Subsequently, hepatic cholesterol 7α-hydroxylase (CYP7A1) and oxysterol 7α-hydroxylase (CYP7B1) were inhibited. Moreover, the two strains enhanced BA transport by increasing the levels of hepatic multidrug resistance-associated protein homologs 3 and 4 (Mrp3/4), hepatic multidrug resistance protein 2 (Mdr2), and the bile salt export pump (BSEP). In addition, both L. reuteri and L. plantarum reduced LD-associated gut microbiota dysbiosis. L. reuteri increased the relative abundance of Muribaculaceae, while L. plantarum increased that of Akkermansia. The changed gut microbiota was significantly negatively correlated with the incidence of cholesterol gallstones and the FXR-antagonistic BAs in the liver and serum and with the FXR signaling pathways. Furthermore, the protective effects of the two strains were abolished by both global and intestine-specific FXR antagonists. These findings suggest that Lactobacillus might relieve CGS through the FXR signaling pathways. IMPORTANCE Cholesterol gallstone (CGS) disease is prevalent worldwide. None of the medical options for prevention and treatment of CGS disease are recommended, and surgical management has a high rate of recurrence. It has been reported that the factors involved in metabolic syndrome are highly connected with CGS formation. While remodeling of dysbiosis of the gut microbiome during improvement of metabolic syndrome has been well studied, less is known about prevention of CGS formation after regulating the gut microbiome. We used the lithogenic diet (LD) to induce an experimental CGS model in C57BL/6J mice to investigate protection against CGS formation by Limosilactobacillus reuteri strain CGMCC 17942 and Lactiplantibacillus plantarum strain CGMCC 14407. We found that these L. reuteri and L. plantarum strains altered the bile acid composition in mice and improved the dysbiosis of the gut microbiome. These two Lactobacillus strains prevented CGS formation by fully activating the hepatic and ileal FXR signaling pathways. They could be a promising therapeutic strategy for treating CGS or preventing its recurrence.
Keywords: FGF15; FXR; Lactobacillus; bile acid; cholesterol gallstones; gut microbiota.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
