

CircRNA-CREIT inhibits stress granule assembly and overcomes doxorubicin resistance in TNBC by destabilizing PKR
- PMID: 36038948
- PMCID: PMC9425971
- DOI: 10.1186/s13045-022-01345-w
CircRNA-CREIT inhibits stress granule assembly and overcomes doxorubicin resistance in TNBC by destabilizing PKR
Abstract
Background: Circular RNAs (circRNAs) represent a novel type of regulatory RNA characterized by high evolutionary conservation and stability. CircRNAs are expected to be potential diagnostic biomarkers and therapeutic targets for a variety of malignancies. However, the regulatory functions and underlying mechanisms of circRNAs in triple-negative breast cancer (TNBC) are largely unknown.
Methods: By using RNA high-throughput sequencing technology, qRT-PCR and in situ hybridization assays, we screened dysregulated circRNAs in breast cancer and TNBC tissues. Then in vitro assays, animal models and patient-derived organoids (PDOs) were utilized to explore the roles of the candidate circRNA in TNBC. To investigate the underlying mechanisms, RNA pull-down, RNA immunoprecipitation (RIP), co immunoprecipitation (co-IP) and Western blotting assays were carried out.
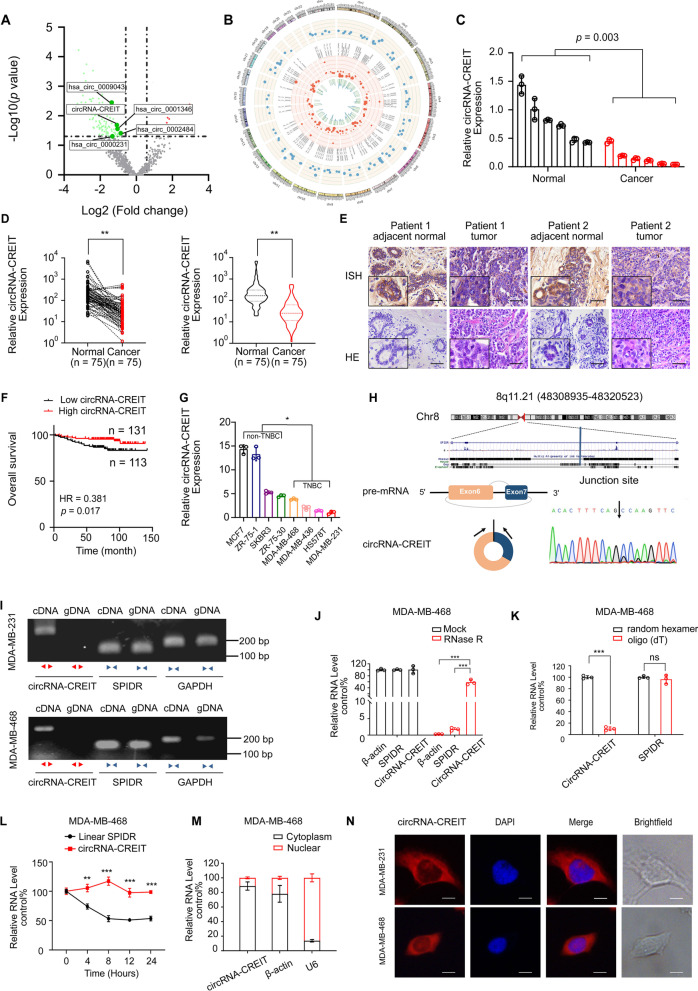
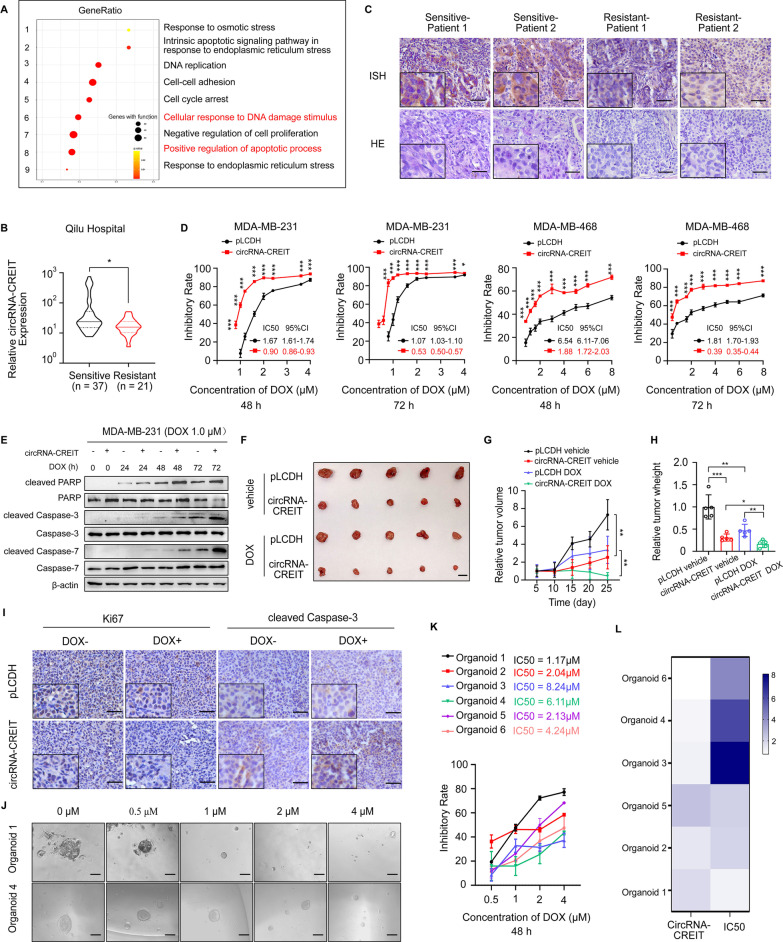
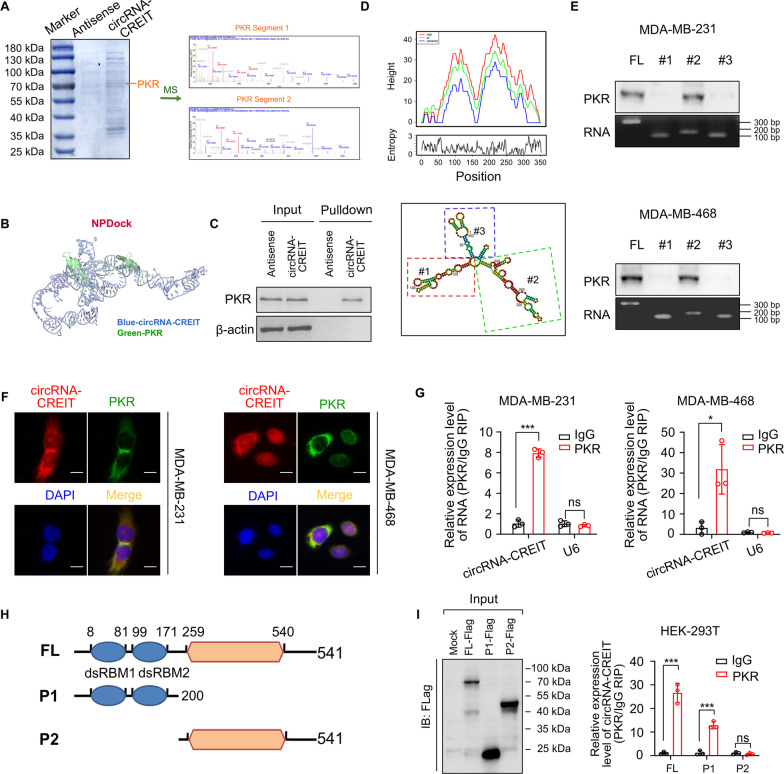
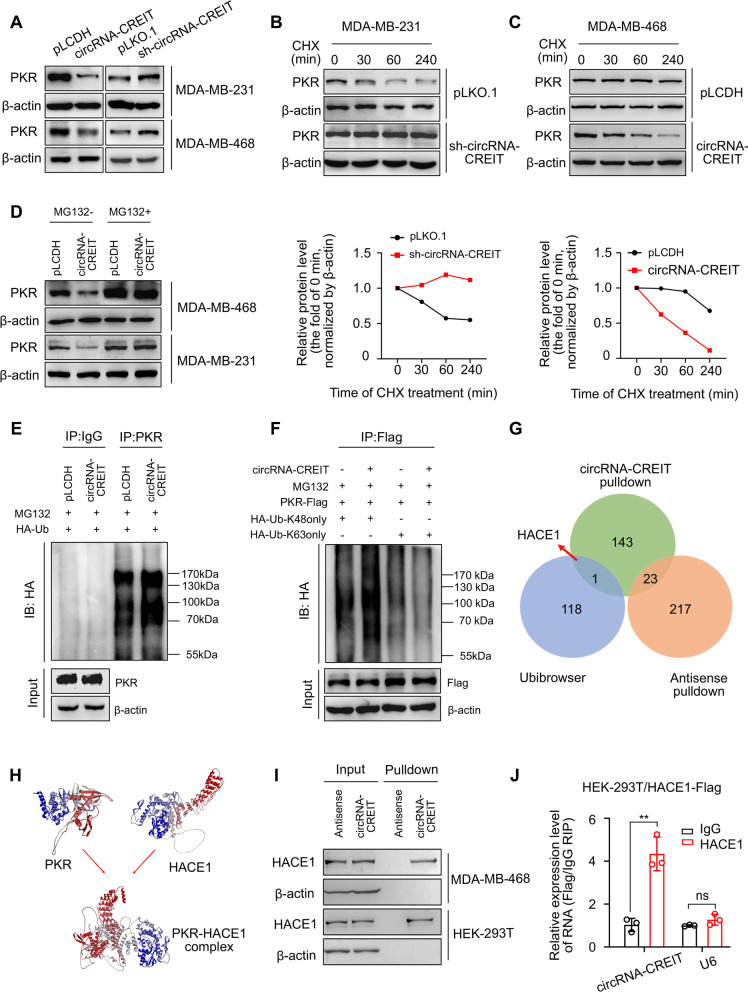
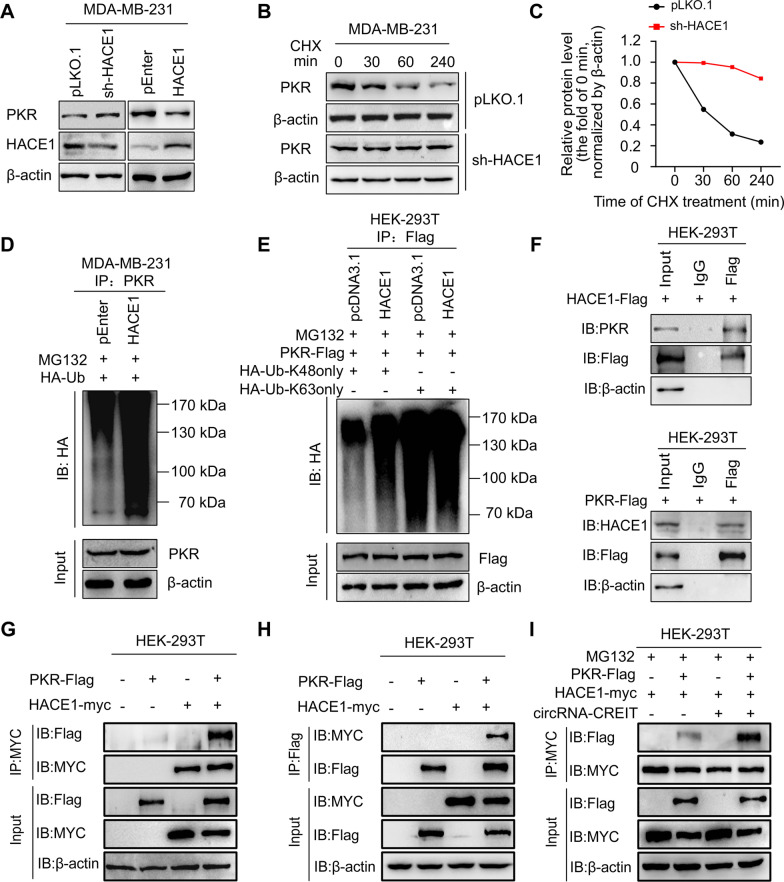
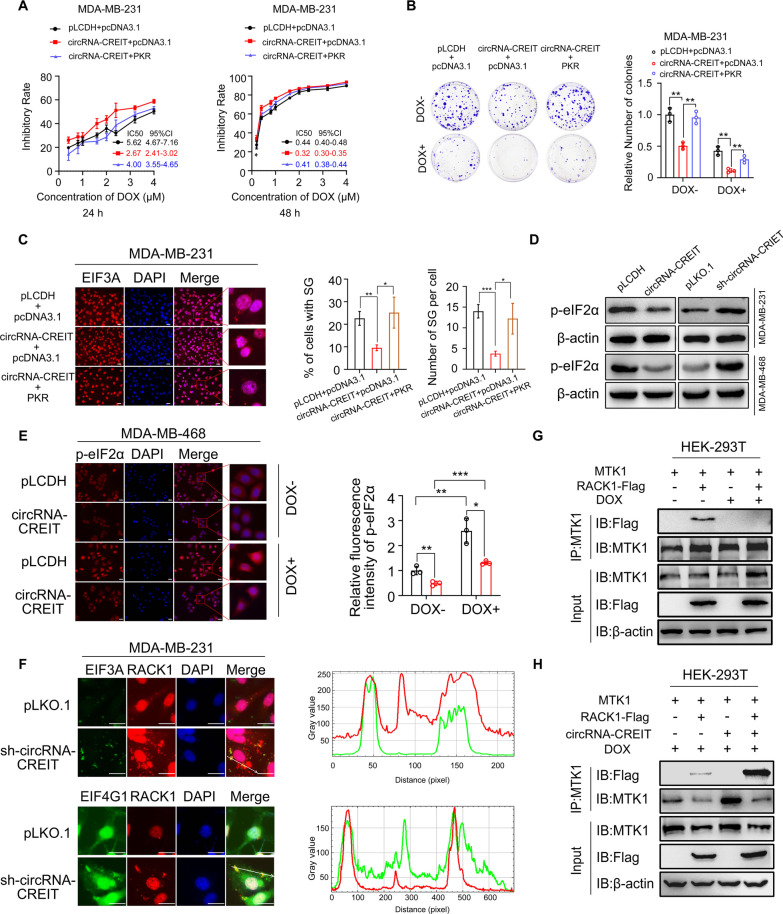
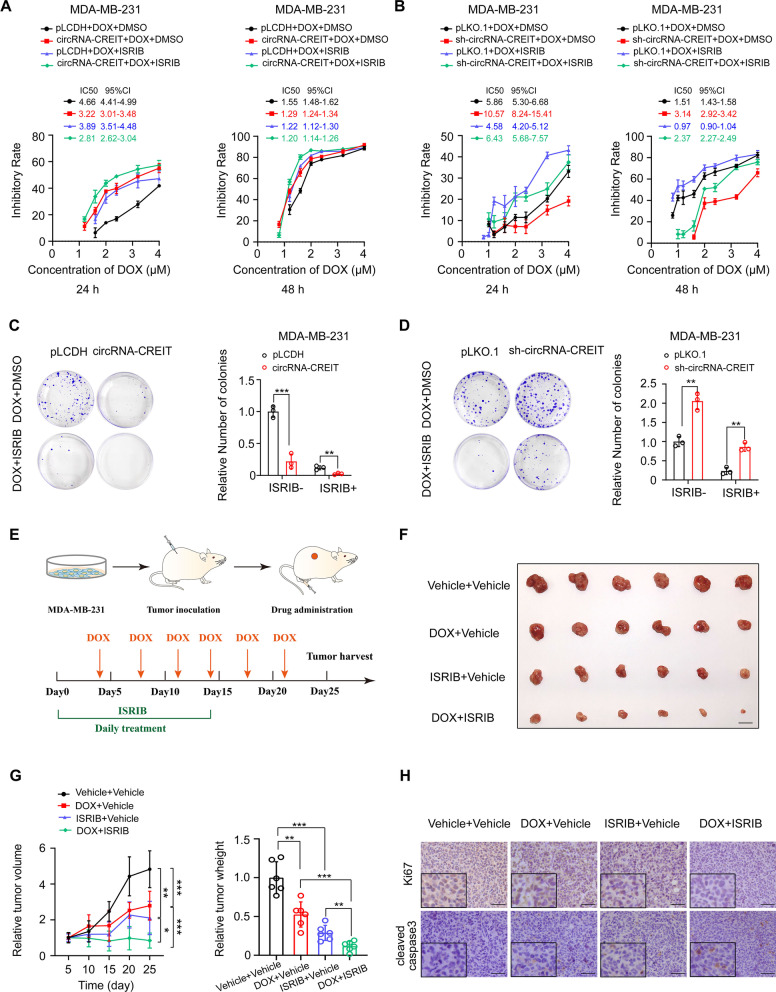
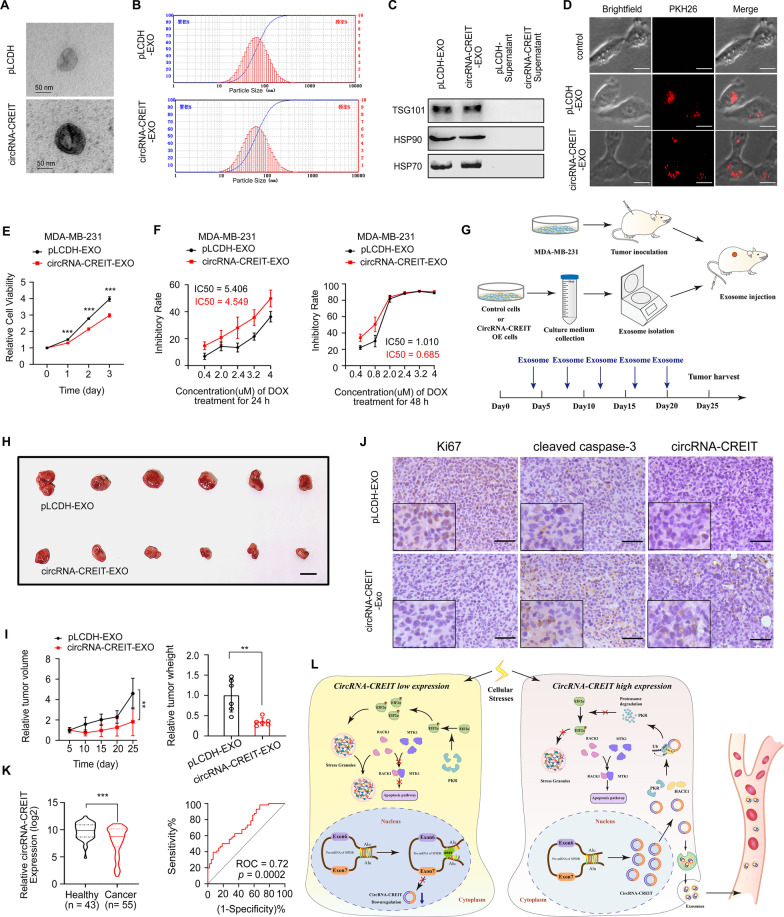
Results: In this study, we demonstrated that circRNA-CREIT was aberrantly downregulated in doxorubicin resistant triple-negative breast cancer (TNBC) cells and associated with a poor prognosis. The RNA binding protein DHX9 was responsible for the reduction in circRNA-CREIT by interacting with the flanking inverted repeat Alu (IRAlu) sequences and inhibiting back-splicing. By utilizing in vitro assays, animal models and patient-derived organoids, we revealed that circRNA-CREIT overexpression significantly enhanced the doxorubicin sensitivity of TNBC cells. Mechanistically, circRNA-CREIT acted as a scaffold to facilitate the interaction between PKR and the E3 ligase HACE1 and promoted proteasomal degradation of PKR protein via K48-linked polyubiquitylation. A reduced PKR/eIF2α signaling axis was identified as a critical downstream effector of circRNA-CREIT, which attenuated the assembly of stress granules (SGs) to activate the RACK1/MTK1 apoptosis signaling pathway. Further investigations revealed that a combination of the SG inhibitor ISRIB and doxorubicin synergistically inhibited TNBC tumor growth. Besides, circRNA-CREIT could be packaged into exosomes and disseminate doxorubicin sensitivity among TNBC cells.
Conclusions: Our study demonstrated that targeting circRNA-CREIT and SGs could serve as promising therapeutic strategies against TNBC chemoresistance.
Keywords: Chemoresistance; CircRNA-CREIT; Stress granules; TNBC.
© 2022. The Author(s).
Conflict of interest statement
All the authors declare they have no competing interests.
Figures
References
-
- Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomataram I, Jemal A, et al. Global Cancer Statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA 2021;71(3):209–49. - PubMed
-
- Zhu Y, Zhu X, Tang C, Guan X, Zhang W. Progress and challenges of immunotherapy in triple-negative breast cancer. Biochim Biophys Acta. 2021;1876(2):188593. - PubMed
-
- Zhu Y, Hu Y, Tang C, Guan X, Zhang W. Platinum-based systematic therapy in triple-negative breast cancer. Biochim Biophys Acta. 2022;1877(1):188678. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- 82004122/National Natural Science Foundation of China
- 81874119/National Natural Science Foundation of China
- 2020M682199/China Postdoctoral Science Foundation
- ZR2020QH335/Shandong Provincial Natural Science Foundation, China
- CXPJJH121001-2021003/Chen Xiao-ping Foundation for the Development of Science and Technology of Hubei Province
- YXH2022ZX02160/2021 Shandong Medical Association Clinical Research Fund
- 2020YFA0712400/National Key Research and Development Program
- ts20190971/Special Foundation for Taishan Scholars
- 2020SDUCRCA015/Foundation from Clinical Research Center of Shandong University
- 2019-3/Qilu Hospital Clinical New Technology Developing Foundation
LinkOut - more resources
Full Text Sources
Miscellaneous
