

Lysine 27 dimethylation of Drosophila linker histone dH1 contributes to heterochromatin organization independently of H3K9 methylation
- PMID: 36039761
- PMCID: PMC9458452
- DOI: 10.1093/nar/gkac716
Lysine 27 dimethylation of Drosophila linker histone dH1 contributes to heterochromatin organization independently of H3K9 methylation
Abstract
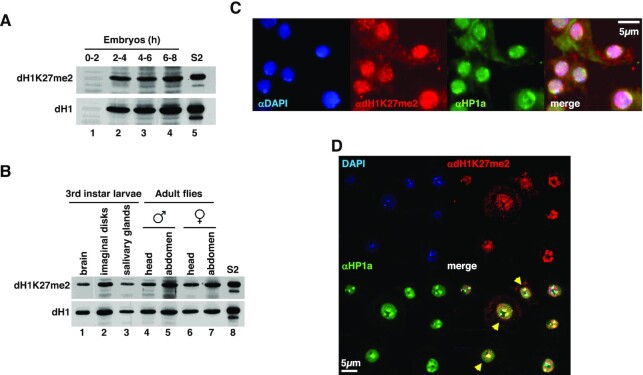
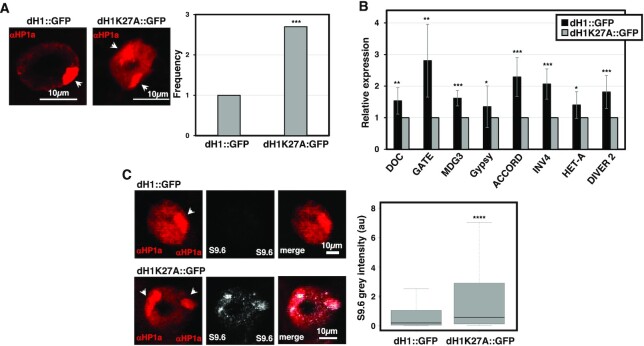
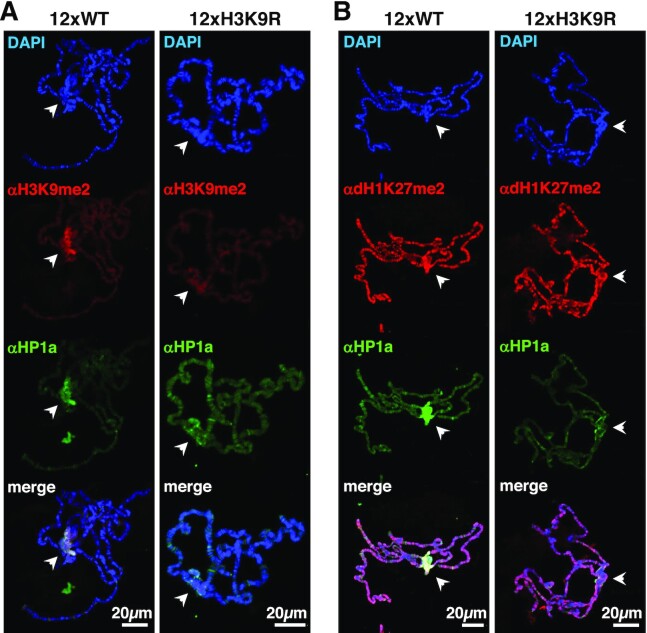
Post-translational modifications (PTMs) of core histones are important epigenetic determinants that correlate with functional chromatin states. However, despite multiple linker histone H1s PTMs have been identified, little is known about their genomic distribution and contribution to the epigenetic regulation of chromatin. Here, we address this question in Drosophila that encodes a single somatic linker histone, dH1. We previously reported that dH1 is dimethylated at K27 (dH1K27me2). Here, we show that dH1K27me2 is a major PTM of Drosophila heterochromatin. At mitosis, dH1K27me2 accumulates at pericentromeric heterochromatin, while, in interphase, it is also detected at intercalary heterochromatin. ChIPseq experiments show that >98% of dH1K27me2 enriched regions map to heterochromatic repetitive DNA elements, including transposable elements, simple DNA repeats and satellite DNAs. Moreover, expression of a mutated dH1K27A form, which impairs dH1K27me2, alters heterochromatin organization, upregulates expression of heterochromatic transposable elements and results in the accumulation of RNA:DNA hybrids (R-loops) in heterochromatin, without affecting H3K9 methylation and HP1a binding. The pattern of dH1K27me2 is H3K9 methylation independent, as it is equally detected in flies carrying a H3K9R mutation, and is not affected by depletion of Su(var)3-9, HP1a or Su(var)4-20. Altogether these results suggest that dH1K27me2 contributes to heterochromatin organization independently of H3K9 methylation.
© The Author(s) 2022. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures





References
-
- Kouzarides T. Chromatin modifications and their function. Cell. 2007; 128:693–705. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
