

Keratin 8 is a scaffolding and regulatory protein of ERAD complexes
- PMID: 36045259
- PMCID: PMC11803007
- DOI: 10.1007/s00018-022-04528-3
Keratin 8 is a scaffolding and regulatory protein of ERAD complexes
Abstract
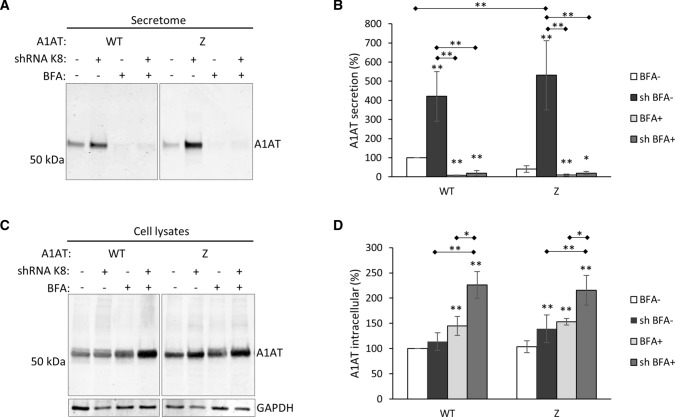
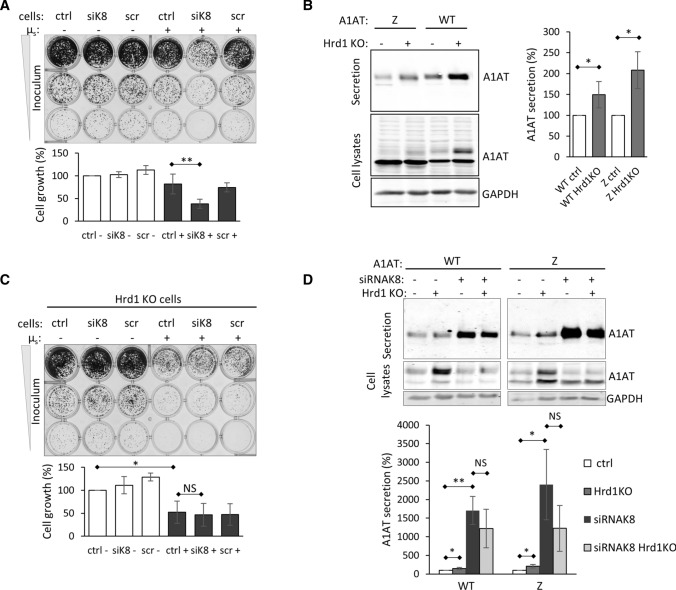
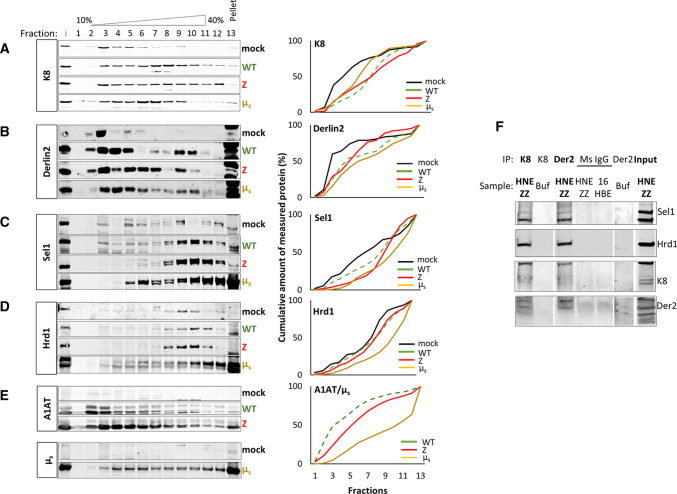
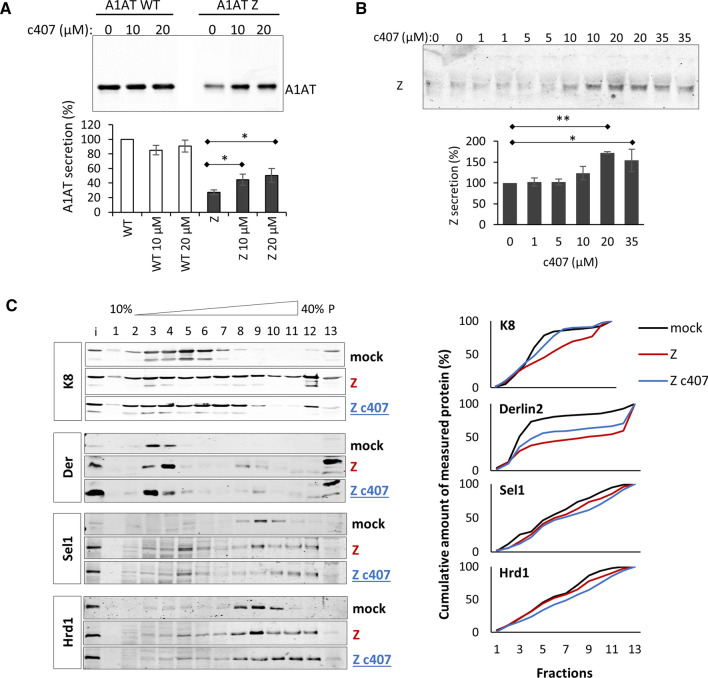
Early recognition and enhanced degradation of misfolded proteins by the endoplasmic reticulum (ER) quality control and ER-associated degradation (ERAD) cause defective protein secretion and membrane targeting, as exemplified for Z-alpha-1-antitrypsin (Z-A1AT), responsible for alpha-1-antitrypsin deficiency (A1ATD) and F508del-CFTR (cystic fibrosis transmembrane conductance regulator) responsible for cystic fibrosis (CF). Prompted by our previous observation that decreasing Keratin 8 (K8) expression increased trafficking of F508del-CFTR to the plasma membrane, we investigated whether K8 impacts trafficking of soluble misfolded Z-A1AT protein. The subsequent goal of this study was to elucidate the mechanism underlying the K8-dependent regulation of protein trafficking, focusing on the ERAD pathway. The results show that diminishing K8 concentration in HeLa cells enhances secretion of both Z-A1AT and wild-type (WT) A1AT with a 13-fold and fourfold increase, respectively. K8 down-regulation triggers ER failure and cellular apoptosis when ER stress is jointly elicited by conditional expression of the µs heavy chains, as previously shown for Hrd1 knock-out. Simultaneous K8 silencing and Hrd1 knock-out did not show any synergistic effect, consistent with K8 acting in the Hrd1-governed ERAD step. Fractionation and co-immunoprecipitation experiments reveal that K8 is recruited to ERAD complexes containing Derlin2, Sel1 and Hrd1 proteins upon expression of Z/WT-A1AT and F508del-CFTR. Treatment of the cells with c407, a small molecule inhibiting K8 interaction, decreases K8 and Derlin2 recruitment to high-order ERAD complexes. This was associated with increased Z-A1AT secretion in both HeLa and Z-homozygous A1ATD patients' respiratory cells. Overall, we provide evidence that K8 acts as an ERAD modulator. It may play a scaffolding protein role for early-stage ERAD complexes, regulating Hrd1-governed retrotranslocation initiation/ubiquitination processes. Targeting K8-containing ERAD complexes is an attractive strategy for the pharmacotherapy of A1ATD.
Keywords: Cytoskeleton; Epithelium; Intermediary filaments; Protein complexes fractionation; Protein–protein interaction; Synthetic lethality.
© 2022. The Author(s), under exclusive licence to Springer Nature Switzerland AG.
Conflict of interest statement
The authors have no relevant financial or non-financial interests to disclose.
Figures



References
-
- Chakraborty P, Teckman J (2014) Alpha-1-antitrypsin deficiency liver disease: science and therapeutic potential 50 years later. J Gastroenterol Pancreatol Liver Disord 1(3):1–9. 10.15226/2374-815X/1/3/00113 - DOI
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
