

The role of somatosensory innervation of adipose tissues
- PMID: 36045288
- PMCID: PMC9477745
- DOI: 10.1038/s41586-022-05137-7
The role of somatosensory innervation of adipose tissues
Abstract
Adipose tissues communicate with the central nervous system to maintain whole-body energy homeostasis. The mainstream view is that circulating hormones secreted by the fat convey the metabolic state to the brain, which integrates peripheral information and regulates adipocyte function through noradrenergic sympathetic output1. Moreover, somatosensory neurons of the dorsal root ganglia innervate adipose tissue2. However, the lack of genetic tools to selectively target these neurons has limited understanding of their physiological importance. Here we developed viral, genetic and imaging strategies to manipulate sensory nerves in an organ-specific manner in mice. This enabled us to visualize the entire axonal projection of dorsal root ganglia from the soma to subcutaneous adipocytes, establishing the anatomical underpinnings of adipose sensory innervation. Functionally, selective sensory ablation in adipose tissue enhanced the lipogenic and thermogenetic transcriptional programs, resulting in an enlarged fat pad, enrichment of beige adipocytes and elevated body temperature under thermoneutral conditions. The sensory-ablation-induced phenotypes required intact sympathetic function. We postulate that beige-fat-innervating sensory neurons modulate adipocyte function by acting as a brake on the sympathetic system. These results reveal an important role of the innervation by dorsal root ganglia of adipose tissues, and could enable future studies to examine the role of sensory innervation of disparate interoceptive systems.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
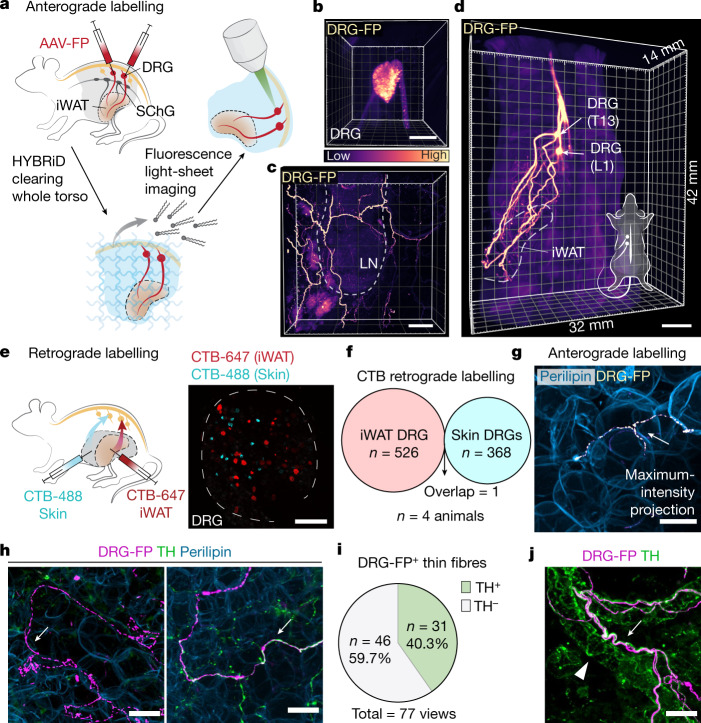
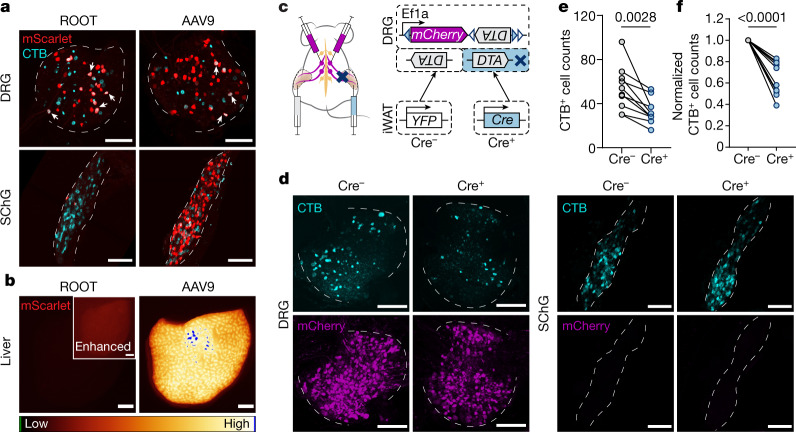
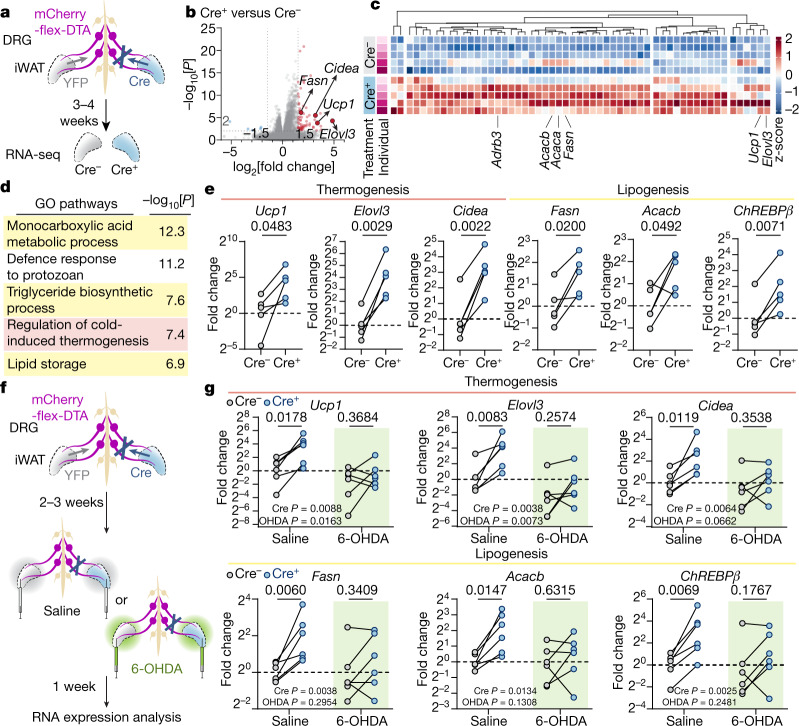
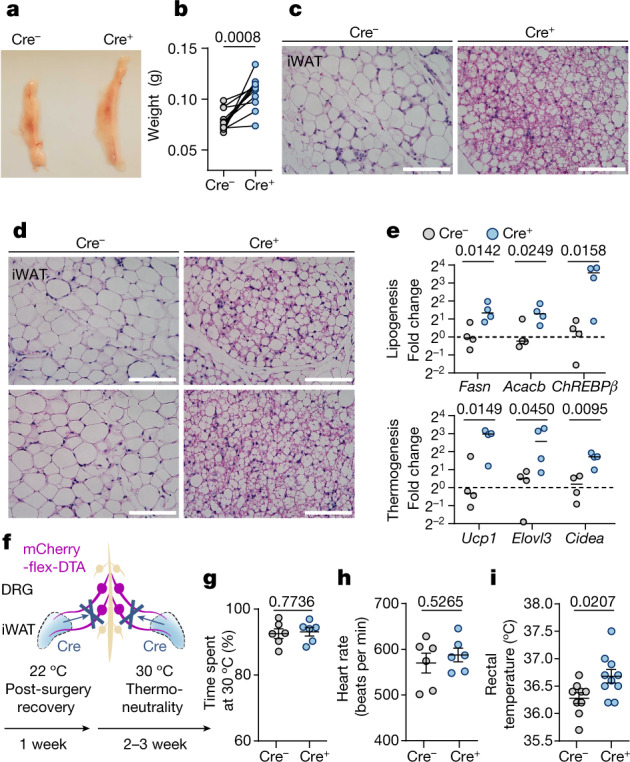
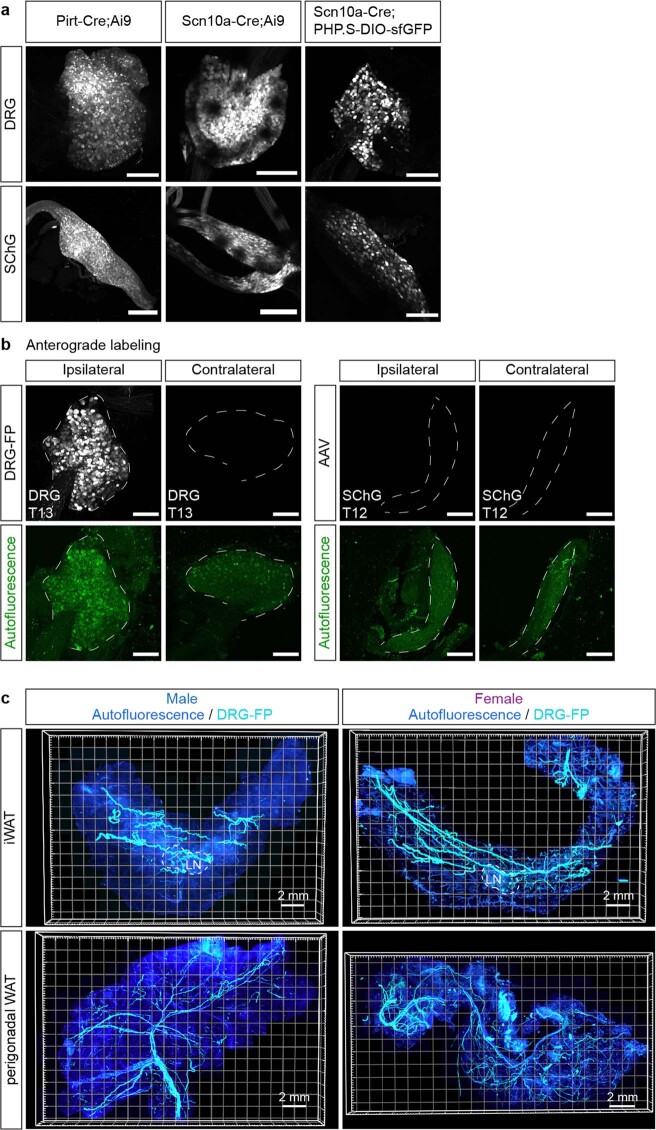
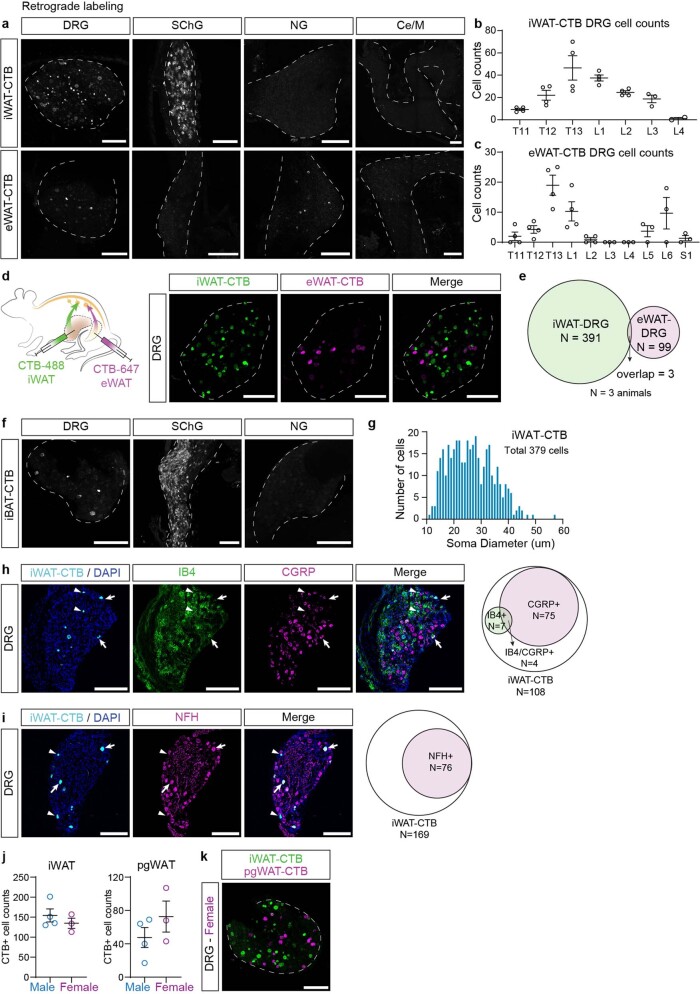
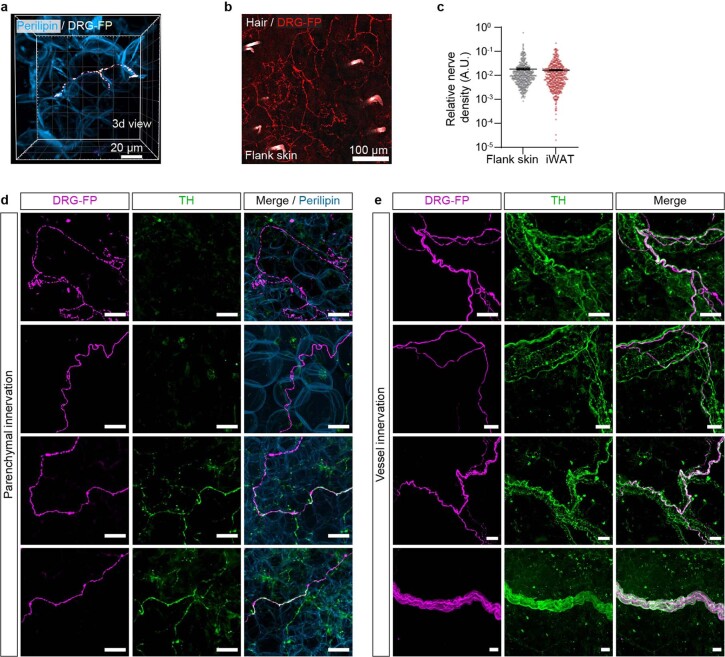
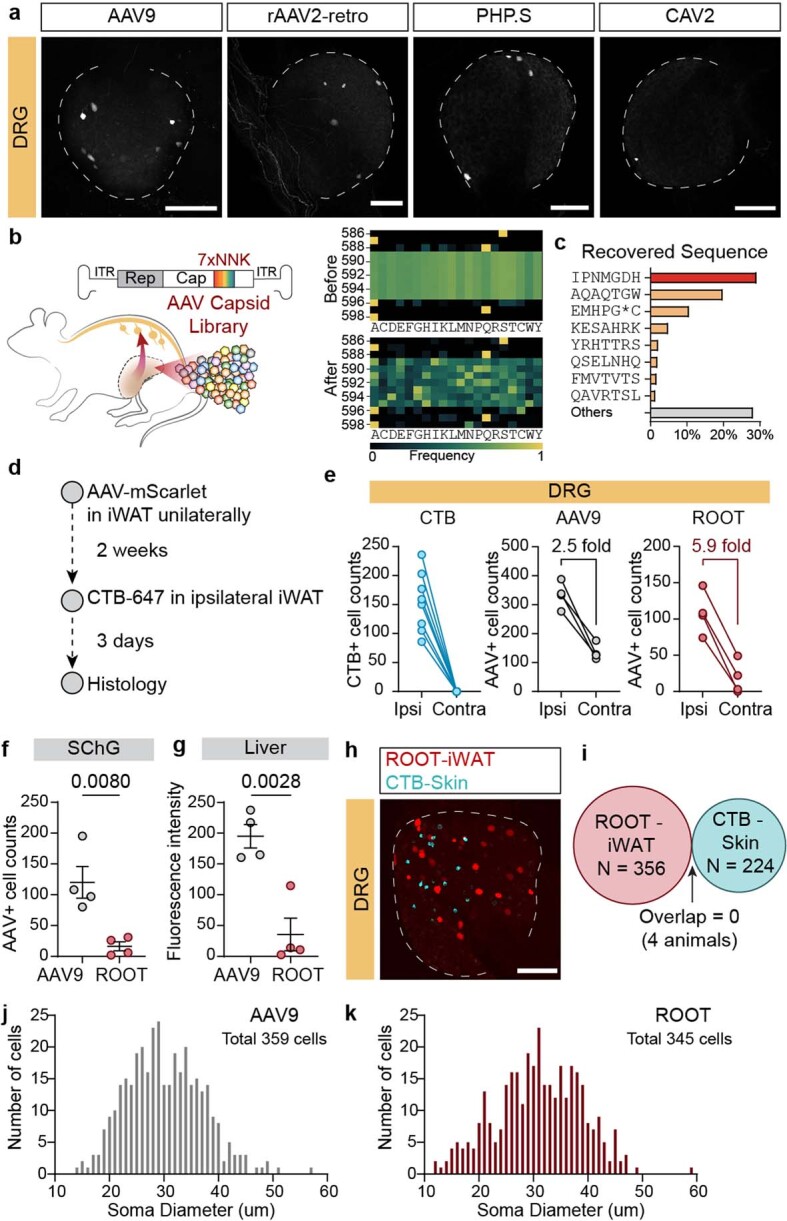
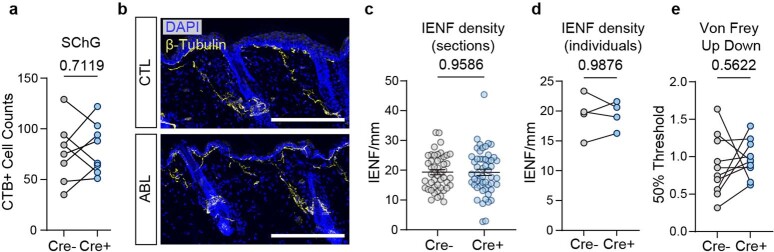
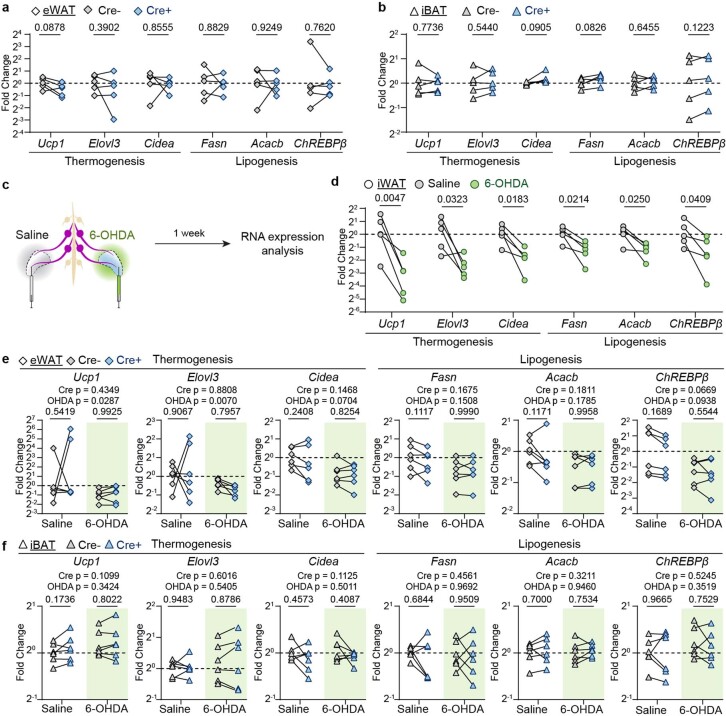
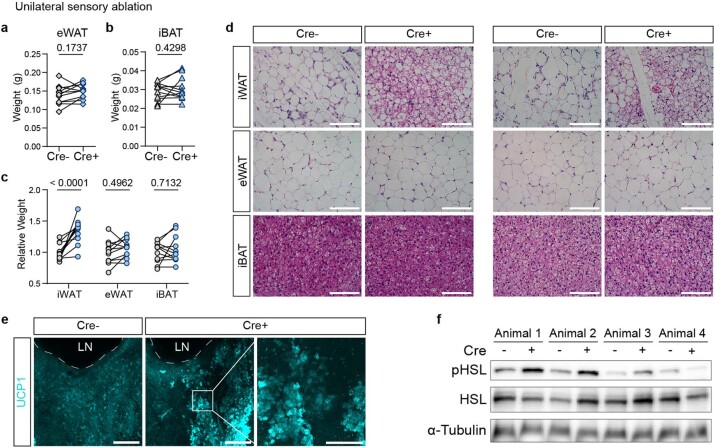
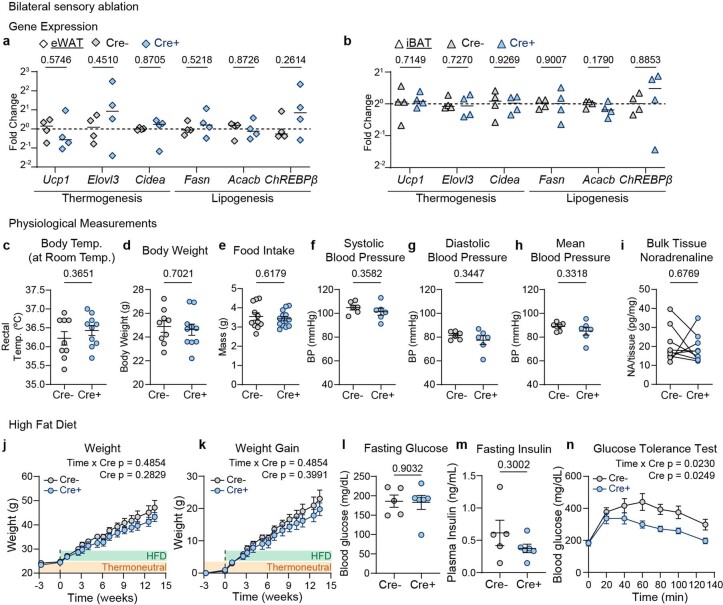
Comment in
-
Sensing and acting: Full circle in fat.Cell Metab. 2023 Jan 3;35(1):9-11. doi: 10.1016/j.cmet.2022.12.010. Cell Metab. 2023. PMID: 36599305
References
-
- Chi J, et al. Three-dimensional adipose tissue imaging reveals regional variation in beige fat biogenesis and PRDM16-dependent sympathetic neurite density. Cell Metab. 2018;27:226–236. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
