

Pla2g2a promotes innate Th2-type immunity lymphocytes to increase B1a cells
- PMID: 36050343
- PMCID: PMC9437038
- DOI: 10.1038/s41598-022-18876-4
Pla2g2a promotes innate Th2-type immunity lymphocytes to increase B1a cells
Abstract
Newborns require early generation of effective innate immunity as a primary physiological mechanism for survival. The neonatal Lin28+Let7- developmental pathway allows increased generation of Th2-type cells and B1a (B-1 B) cells compared to adult cells and long-term maintenance of these initially generated innate cells. For initial B1a cell growth from the neonatal to adult stage, Th2-type IL-5 production from ILC2s and NKT2 cells is important to increase B1a cells. The Th17 increase is dependent on extracellular bacteria, and increased bacteria leads to lower Th2-type generation. Secreted group IIA-phospholipase A2 (sPLA2-IIA) from the Pla2g2a gene can bind to gram-positive bacteria and degrade bacterial membranes, controlling microbiota in the intestine. BALB/c mice are Pla2g2a+, and express high numbers of Th2-type cells and B1a cells. C57BL/6 mice are Pla2g2a-deficient and distinct from the SLAM family, and exhibit fewer NKT2 cells and fewer B1a cells from the neonatal to adult stage. We found that loss of Pla2g2a in the BALB/c background decreased IL-5 from Th2-type ILC2s and NKT2s but increased bacterial-reactive NKT17 cells and MAIT cells, and decreased the number of early-generated B1a cells and MZ B cells and the CD4/CD8 T cell ratio. Low IL-5 by decreased Th2-type cells in Pla2g2a loss led to low early-generated B1a cell growth from the neonatal to adult stage. In anti-thymocyte/Thy-1 autoreactive μκ transgenic (ATAμκ Tg) Pla2g2a+ BALB/c background C.B17 mice generated NKT2 cells that continuously control CD1d+ B1 B cells through old aging and lost CD1d in B1 B cells generating strong B1 ATA B cell leukemia/lymphoma. Pla2g2a-deficient ATAμκTg C57BL/6 mice suppressed the initial B1a cell increase, with low/negative spontaneous leukemia/lymphoma generation. These data confirmed that the presence of Pla2g2a to control bacteria is important to allow the neonatal to adult stage. Pla2g2a promotes innate Th2-type immunity lymphocytes to increase early generated B1a cells.
© 2022. This is a U.S. Government work and not under copyright protection in the US; foreign copyright protection may apply.
Conflict of interest statement
The authors declare no competing interests.
Figures
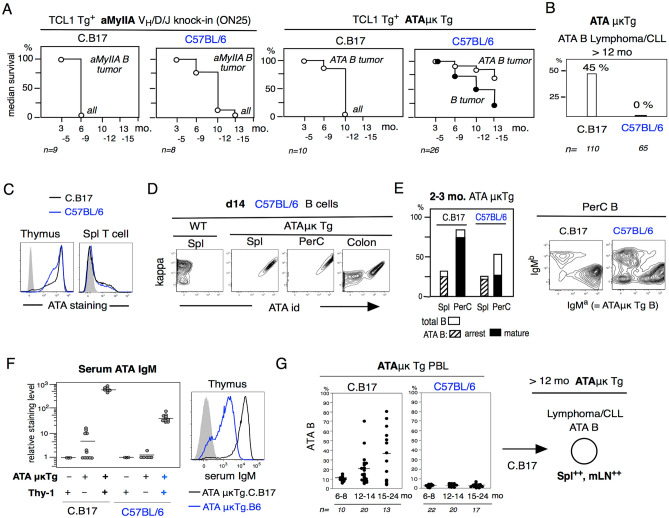



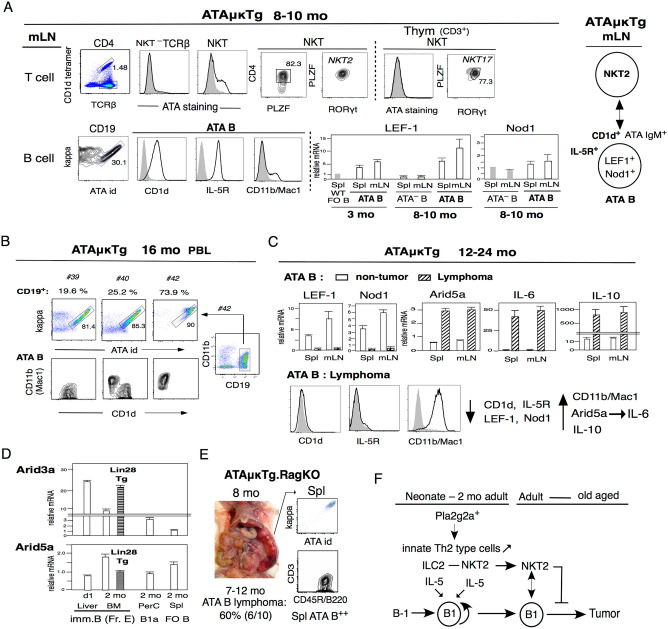
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
