

Rhythmicity of intestinal IgA responses confers oscillatory commensal microbiota mutualism
- PMID: 36054336
- PMCID: PMC7613662
- DOI: 10.1126/sciimmunol.abk2541
Rhythmicity of intestinal IgA responses confers oscillatory commensal microbiota mutualism
Abstract
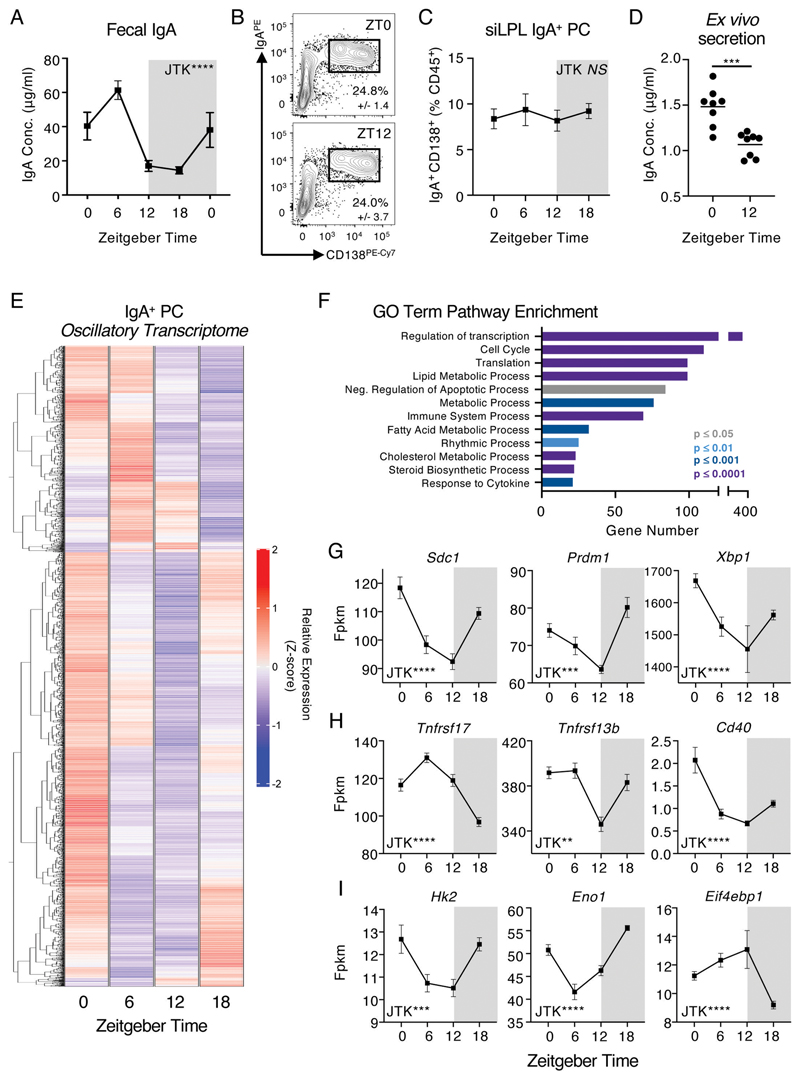
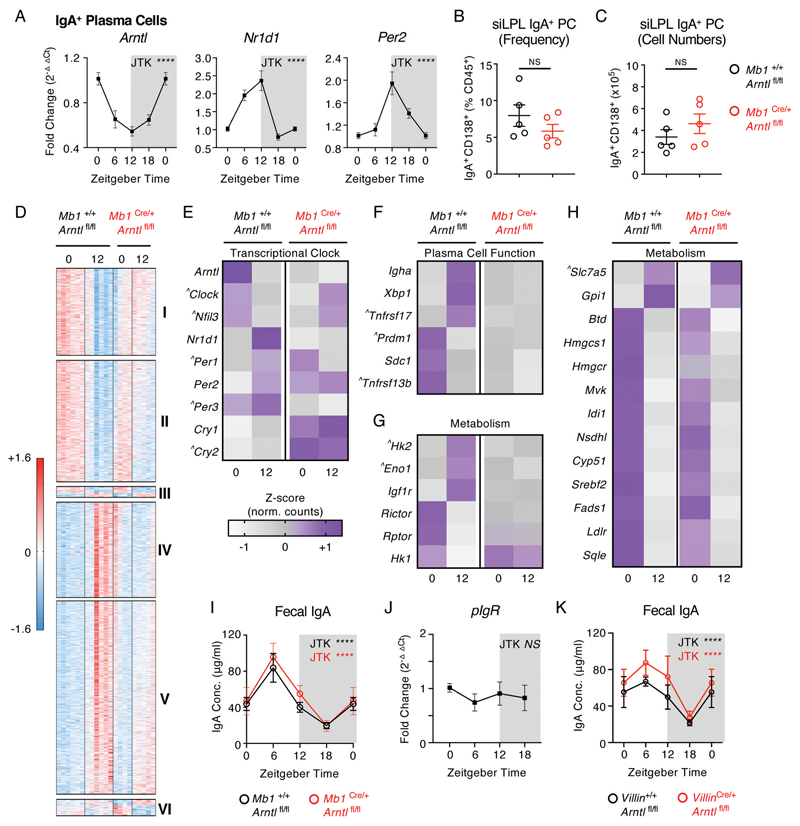
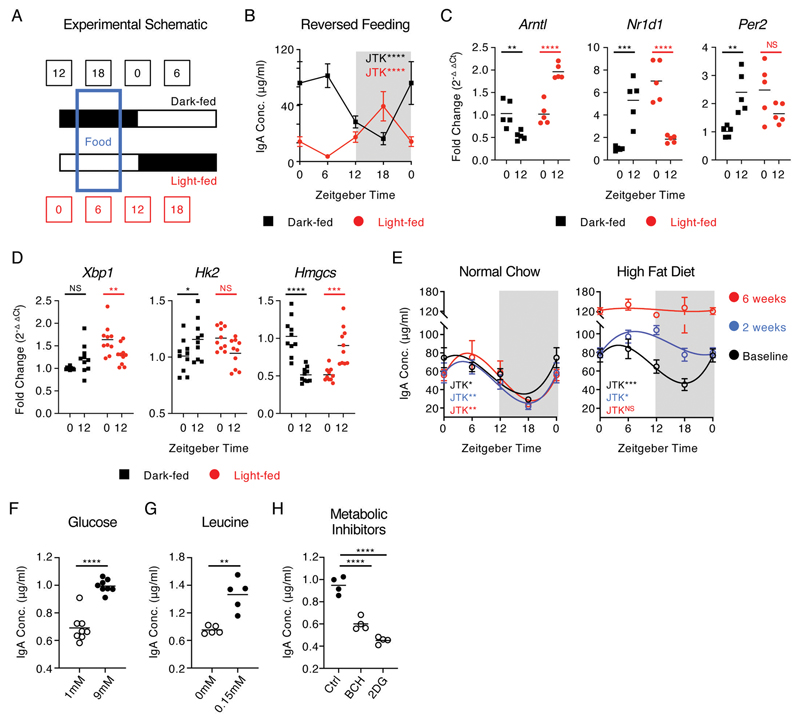
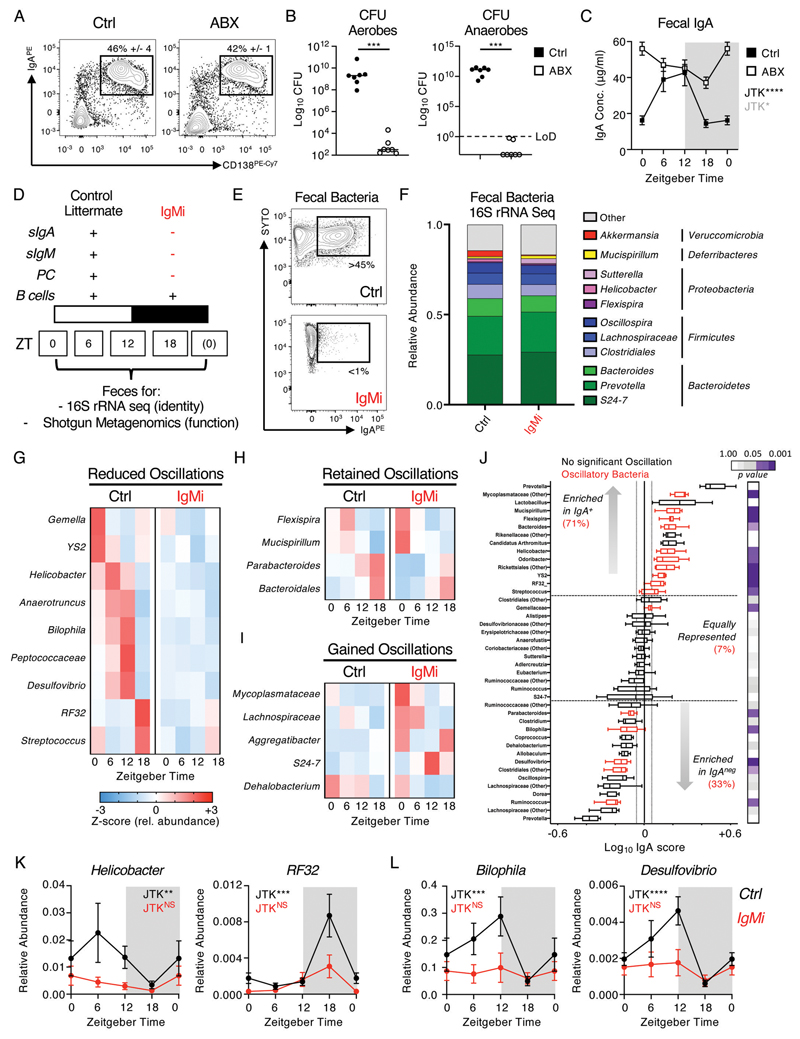
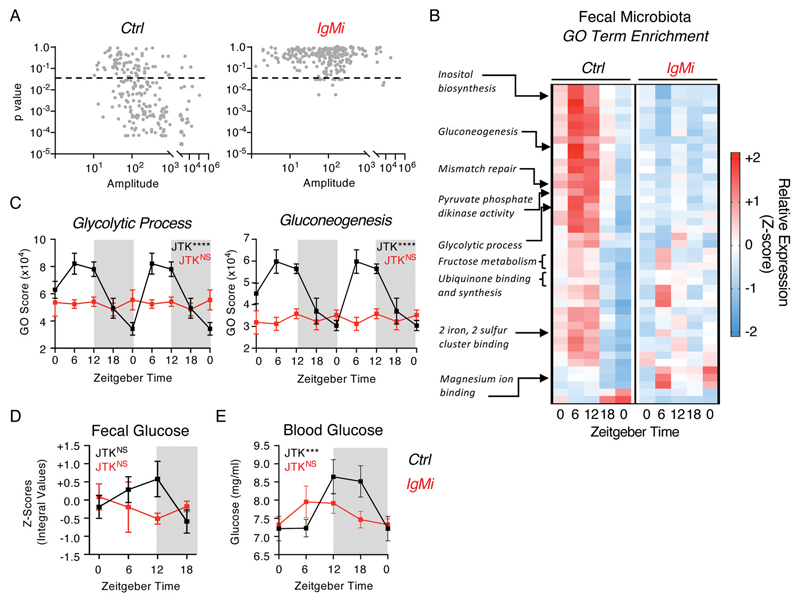
Interactions between the mammalian host and commensal microbiota are enforced through a range of immune responses that confer metabolic benefits and promote tissue health and homeostasis. Immunoglobulin A (IgA) responses directly determine the composition of commensal species that colonize the intestinal tract but require substantial metabolic resources to fuel antibody production by tissue-resident plasma cells. Here, we demonstrate that IgA responses are subject to diurnal regulation over the course of a circadian day. Specifically, the magnitude of IgA secretion, as well as the transcriptome of intestinal IgA+ plasma cells, was found to exhibit rhythmicity. Oscillatory IgA responses were found to be entrained by time of feeding and were also found to be in part coordinated by the plasma cell-intrinsic circadian clock via deletion of the master clock gene Arntl. Moreover, reciprocal interactions between the host and microbiota dictated oscillatory dynamics among the commensal microbial community and its associated transcriptional and metabolic activity in an IgA-dependent manner. Together, our findings suggest that circadian networks comprising intestinal IgA, diet, and the microbiota converge to align circadian biology in the intestinal tract and to ensure host-microbial mutualism.
Conflict of interest statement
Figures
Comment in
-
Feeding IgA+ plasma cells.Nat Rev Immunol. 2022 Nov;22(11):654-655. doi: 10.1038/s41577-022-00788-z. Nat Rev Immunol. 2022. PMID: 36127478 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
- MR/S002715/1/MRC_/Medical Research Council/United Kingdom
- 204796/WT_/Wellcome Trust/United Kingdom
- MR/P00279X/1/MRC_/Medical Research Council/United Kingdom
- MR/N006321/1/MRC_/Medical Research Council/United Kingdom
- 210661/Z/18/Z/WT_/Wellcome Trust/United Kingdom
- G1001390/MRC_/Medical Research Council/United Kingdom
- MR/P001874/1/MRC_/Medical Research Council/United Kingdom
- 097820/WT_/Wellcome Trust/United Kingdom
- BB/T014482/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- 203128/WT_/Wellcome Trust/United Kingdom
- 105644/WT_/Wellcome Trust/United Kingdom
- 203914/Z/16/Z/WT_/Wellcome Trust/United Kingdom
- Z10661/Z/18/Z/WT_/Wellcome Trust/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
