

Low resource availability drives feeding niche partitioning between wild bees and honeybees in a European city
- PMID: 36054537
- PMCID: PMC10077915
- DOI: 10.1002/eap.2727
Low resource availability drives feeding niche partitioning between wild bees and honeybees in a European city
Abstract
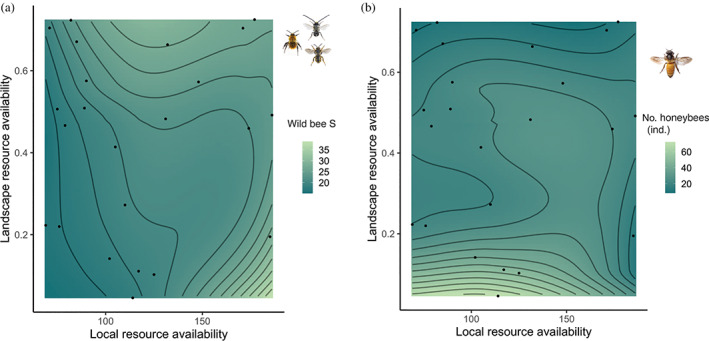
Cities are socioecological systems that filter and select species, therefore establishing unique species assemblages and biotic interactions. Urban ecosystems can host richer wild bee communities than highly intensified agricultural areas, specifically in resource-rich urban green spaces such as allotments and family gardens. At the same time, urban beekeeping has boomed in many European cities, raising concerns that the fast addition of a large number of managed bees could deplete the existing floral resources, triggering competition between wild bees and honeybees. Here, we studied the interplay between resource availability and the number of honeybees at local and landscape scales and how this relationship influences wild bee diversity. We collected wild bees and honeybees in a pollination experiment using four standardized plant species with distinct floral morphologies. We performed the experiment in 23 urban gardens in the city of Zurich (Switzerland), distributed along gradients of urban and local management intensity, and measured functional traits related to resource use. At each site, we quantified the feeding niche partitioning (calculated as the average distance in the multidimensional trait space) between the wild bee community and the honeybee population. Using multilevel structural equation models (SEM), we tested direct and indirect effects of resource availability, urban beekeeping, and wild bees on the community feeding niche partitioning. We found an increase in feeding niche partitioning with increasing wild bee species richness. Moreover, feeding niche partitioning tended to increase in experimental sites with lower resource availability at the landscape scale, which had lower abundances of honeybees. However, beekeeping intensity at the local and landscape scales did not directly influence community feeding niche partitioning or wild bee species richness. In addition, wild bee species richness was positively influenced by local resource availability, whereas local honeybee abundance was positively affected by landscape resource availability. Overall, these results suggest that direct competition for resources was not a main driver of the wild bee community. Due to the key role of resource availability in maintaining a diverse bee community, our study encourages cities to monitor floral resources to better manage urban beekeeping and help support urban pollinators.
Keywords: competition; intraspecific trait variability; pollinator; species interaction; urban biodiversity; urbanization.
© 2022 The Authors. Ecological Applications published by Wiley Periodicals LLC on behalf of The Ecological Society of America.
Conflict of interest statement
The authors declare no conflict of interest.
Figures





References
-
- Albert, C. H. , de Bello F., Boulangeat I., Pellet G., Lavorel S., and Thuiller W.. 2012. “On the Importance of Intraspecific Variability for the Quantification of Functional Diversity.” Oikos 121: 116–26.
-
- Alberti, M. 2015. “Eco‐Evolutionary Dynamics in an Urbanizing Planet.” Trends in Ecology & Evolution 30: 114–26. - PubMed
-
- Baldock, K. C. R. 2020. “Opportunities and Threats for Pollinator Conservation in Global Towns and Cities.” Current Opinion in Insect Science 38: 63–71. - PubMed
-
- Baldock, K. C. R. , Goddard M. A., Hicks D. M., Kunin W. E., Mitschunas N., Osgathorpe L. M., Potts S. G., et al. 2015. “Where is the UK's Pollinator Biodiversity? The Importance of Urban Areas for Flower‐Visiting Insects.” Proceedings of the Royal Society B: Biological Sciences 282(1803): 20142849. 10.1098/rspb.2014.2849. - DOI - PMC - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
