

A serotonergic axon-cilium synapse drives nuclear signaling to alter chromatin accessibility
- PMID: 36055200
- PMCID: PMC9789380
- DOI: 10.1016/j.cell.2022.07.026
A serotonergic axon-cilium synapse drives nuclear signaling to alter chromatin accessibility
Abstract
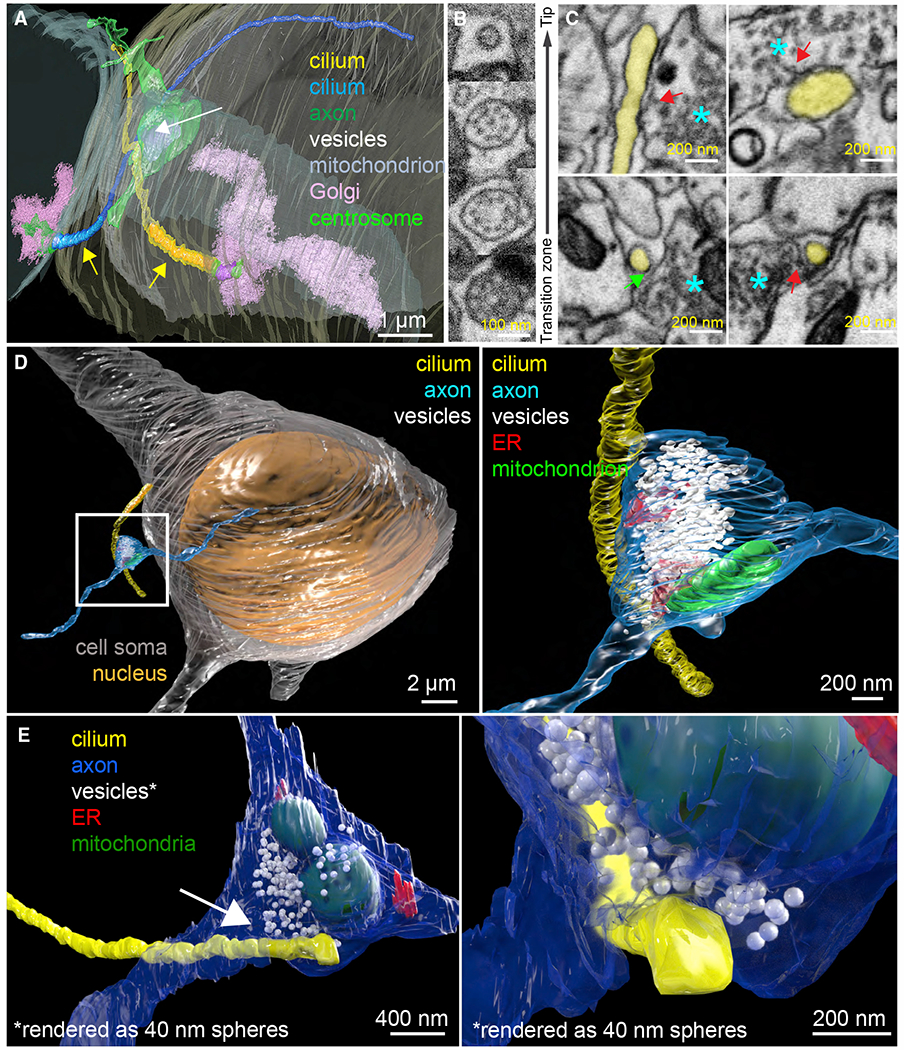
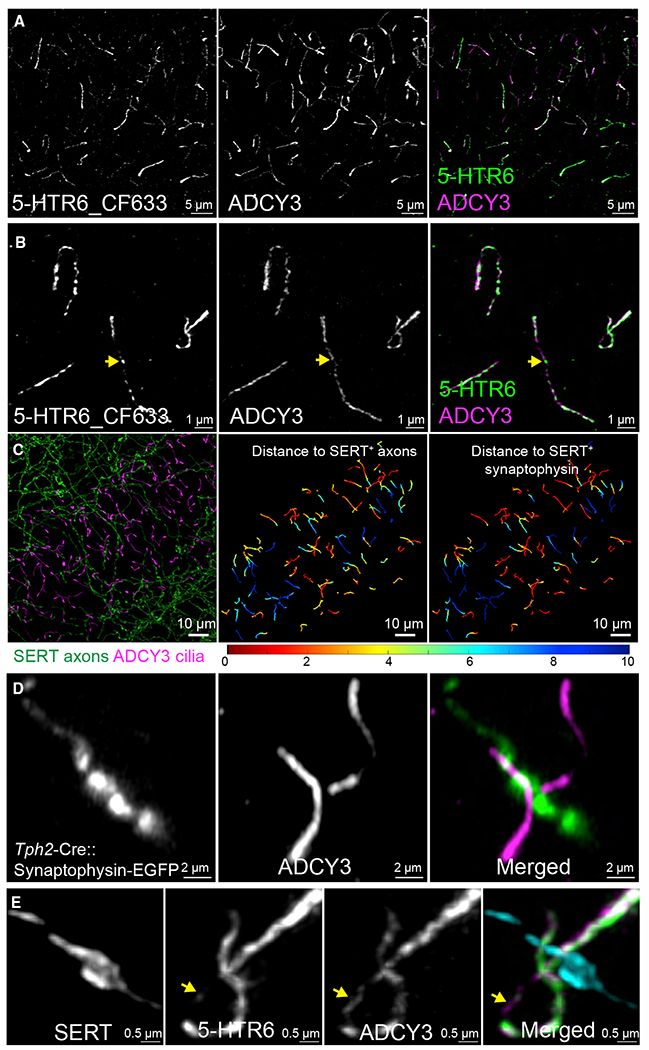
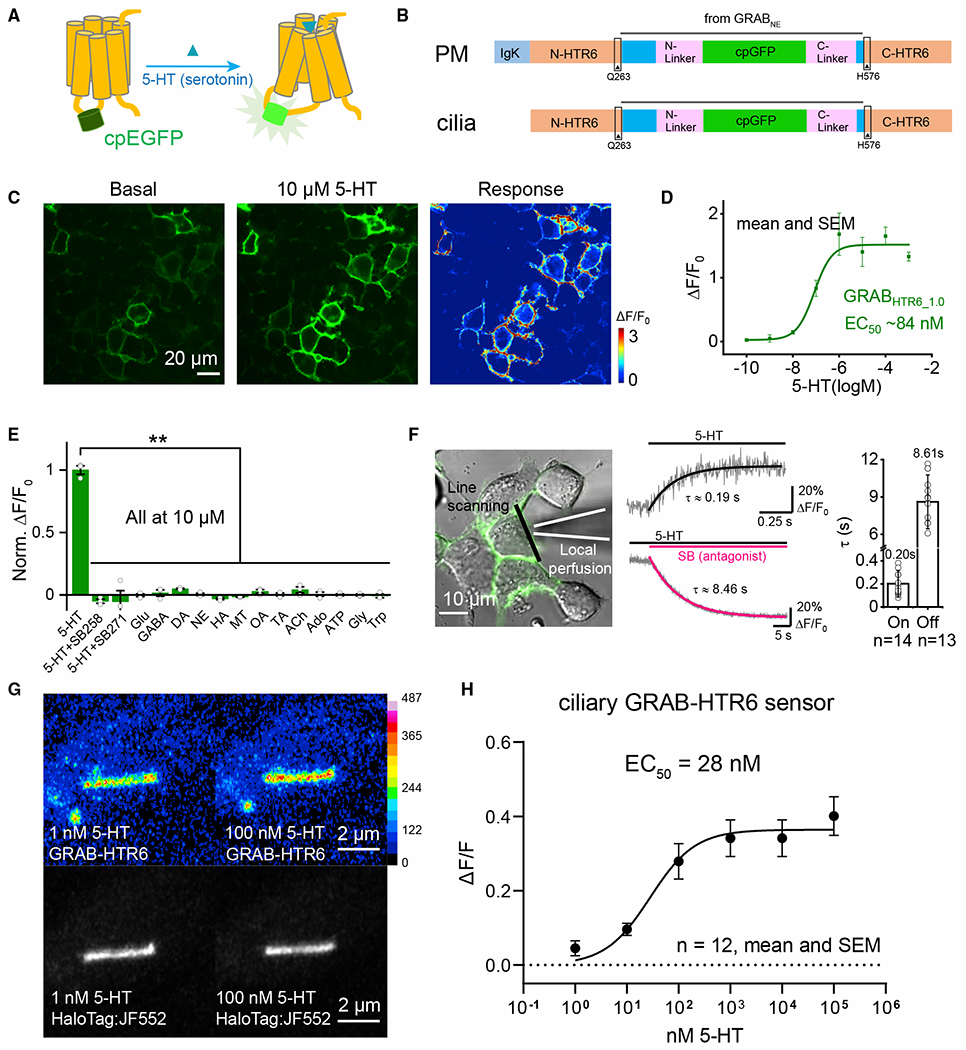
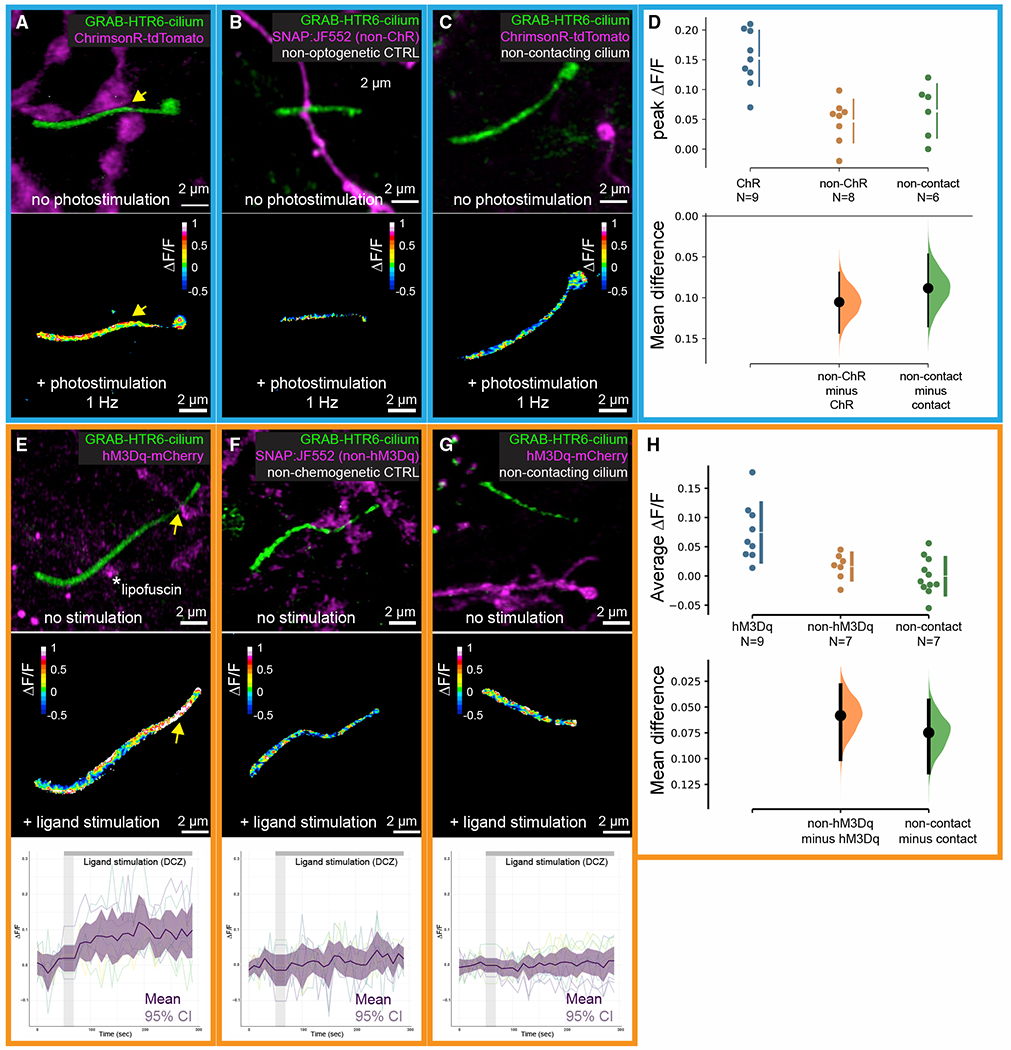
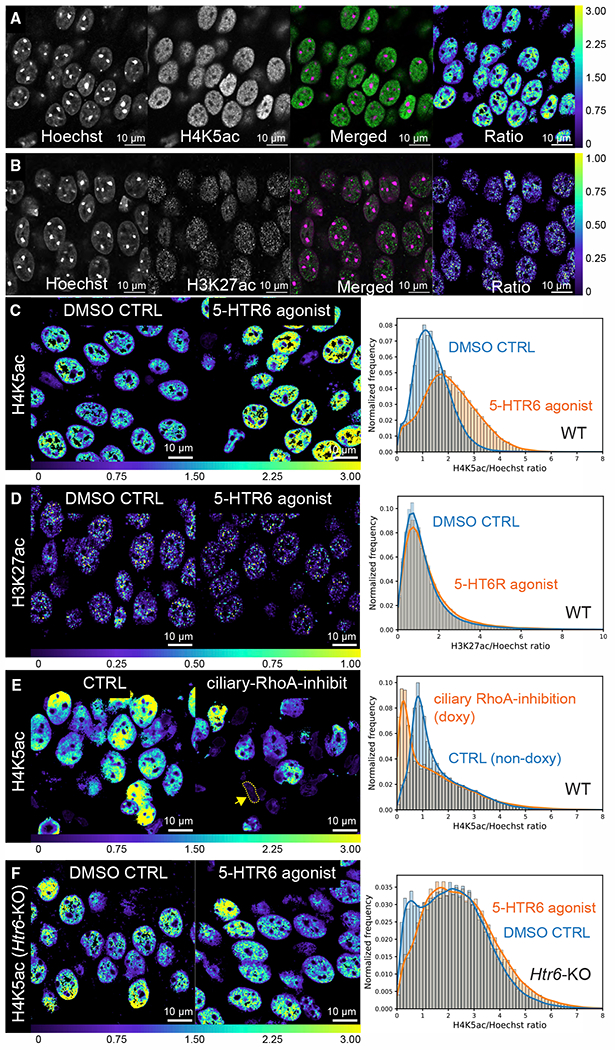
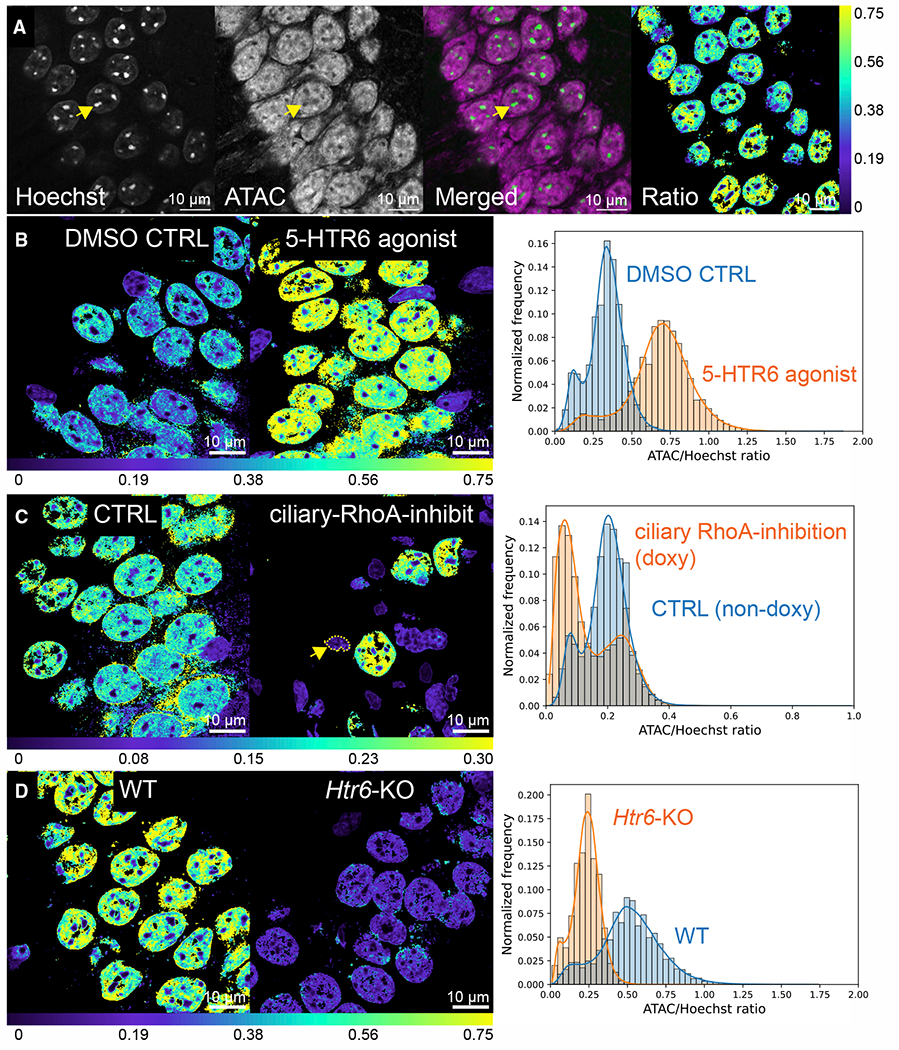
Chemical synapses between axons and dendrites mediate neuronal intercellular communication. Here, we describe a synapse between axons and primary cilia: the axo-ciliary synapse. Using enhanced focused ion beam-scanning electron microscopy on samples with optimally preserved ultrastructure, we discovered synapses between brainstem serotonergic axons and the primary cilia of hippocampal CA1 pyramidal neurons. Functionally, these cilia are enriched in a ciliary-restricted serotonin receptor, the 5-hydroxytryptamine receptor 6 (5-HTR6). Using a cilia-targeted serotonin sensor, we show that opto- and chemogenetic stimulation of serotonergic axons releases serotonin onto cilia. Ciliary 5-HTR6 stimulation activates a non-canonical Gαq/11-RhoA pathway, which modulates nuclear actin and increases histone acetylation and chromatin accessibility. Ablation of this pathway reduces chromatin accessibility in CA1 pyramidal neurons. As a signaling apparatus with proximity to the nucleus, axo-ciliary synapses short circuit neurotransmission to alter the postsynaptic neuron's epigenetic state.
Keywords: FIB-SEM; GPCR signaling; chromatin accessibility; fluorescence lifetime imaging; histone modification; nuclear actin; primary cilia; pyramidal neurons; serotonin.
Copyright © 2022 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests Portions of the technology described herein are covered by U.S. Patent 10,600,615 titled “enhanced FIB-SEM systems for large-volume 3D imaging,” which was issued to C.S.X. and H.F.H. and assigned to Howard Hughes Medical Institute.
Figures

Comment in
-
A ciliary synapse for "short-circuit" neuromodulation.Cell. 2022 Sep 1;185(18):3284-3286. doi: 10.1016/j.cell.2022.08.006. Cell. 2022. PMID: 36055196
-
Cilia make connections.Nat Rev Neurosci. 2022 Nov;23(11):644-645. doi: 10.1038/s41583-022-00636-y. Nat Rev Neurosci. 2022. PMID: 36131051 No abstract available.
References
-
- Alvarez FJ, Pearson JC, Harrington D, Dewey D, Torbeck L, and Fyffe REW (1998). Distribution of 5-hydroxytryptamine-immunoreactive boutons on α-motoneurons in the lumbar spinal cord of adult cats. J. Comp. Neurol 393, 69–83. 10.1002/(sici)1096-9861(19980330)393:1<69::aidcne7>3.0.co;2-o. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
