

Gut bacterial nutrient preferences quantified in vivo
- PMID: 36055202
- PMCID: PMC9450212
- DOI: 10.1016/j.cell.2022.07.020
Gut bacterial nutrient preferences quantified in vivo
Abstract
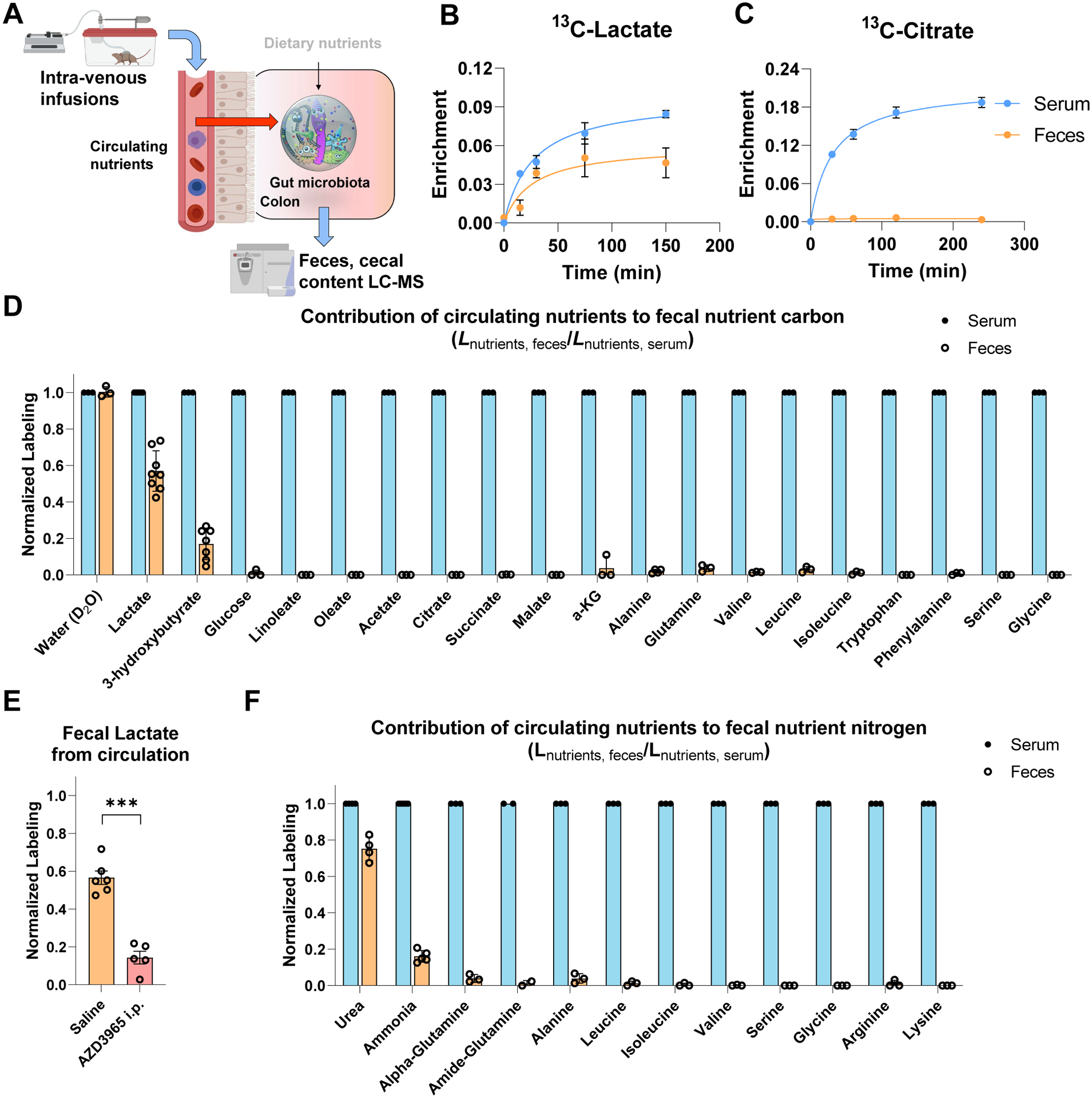
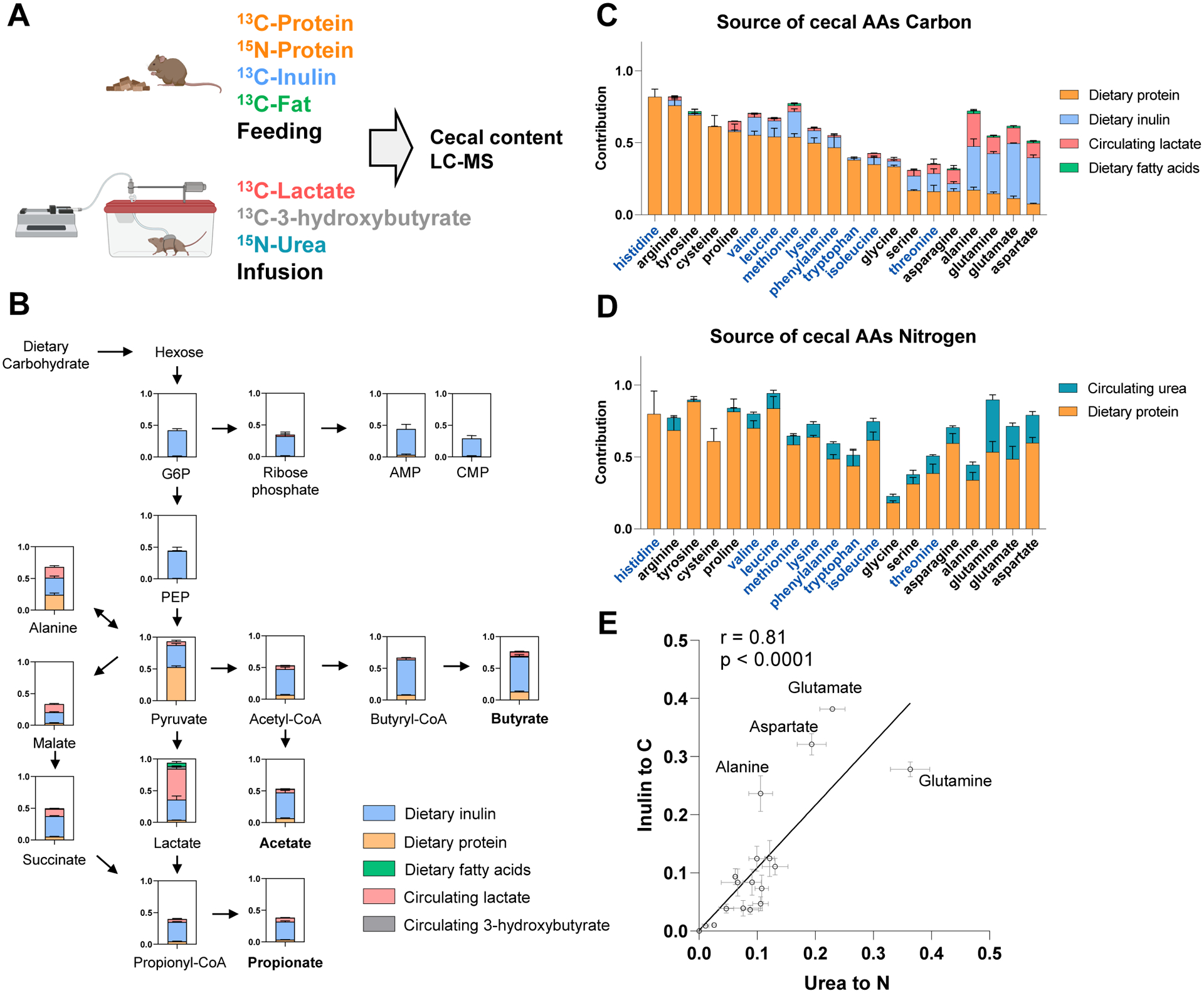
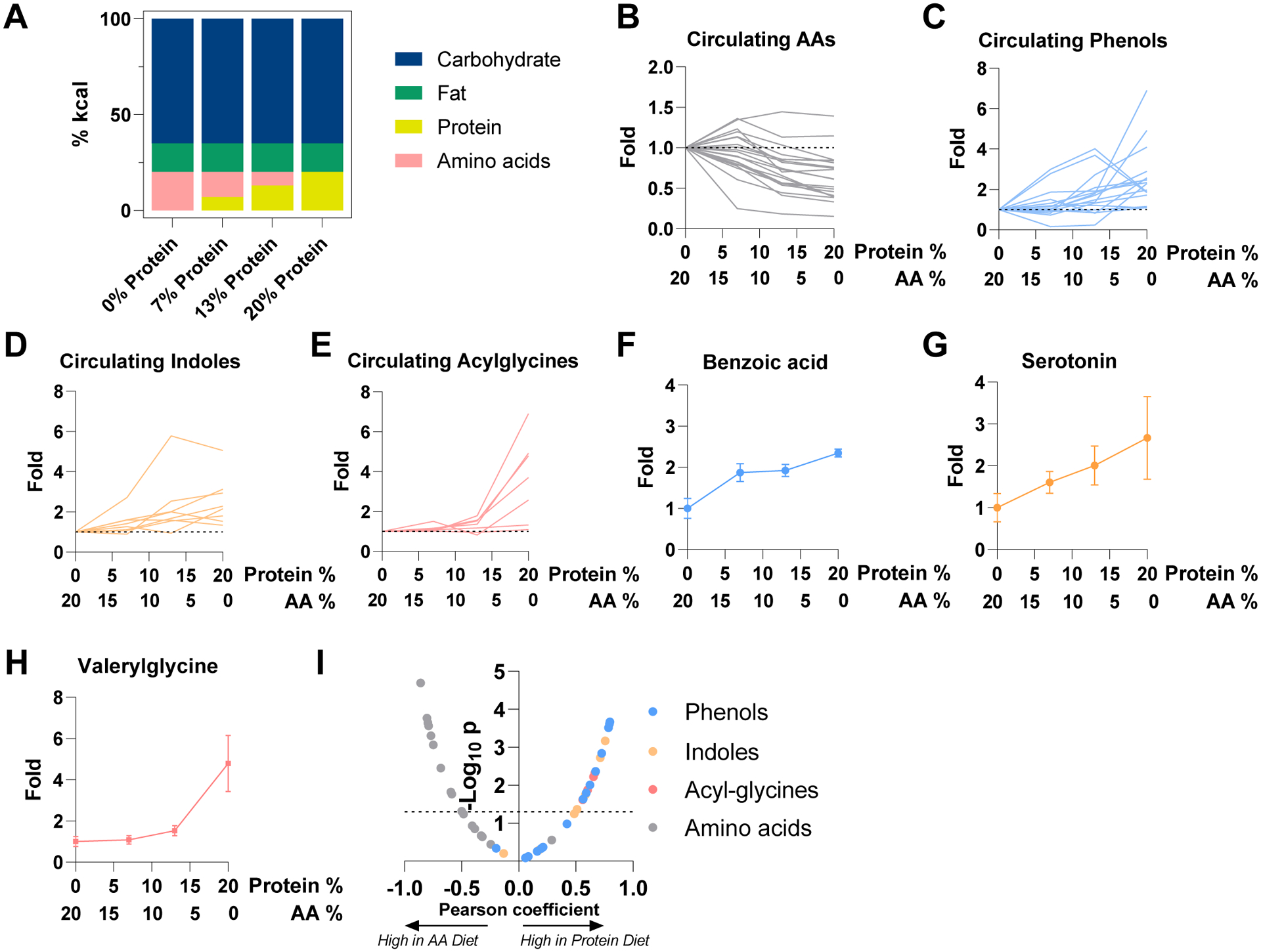
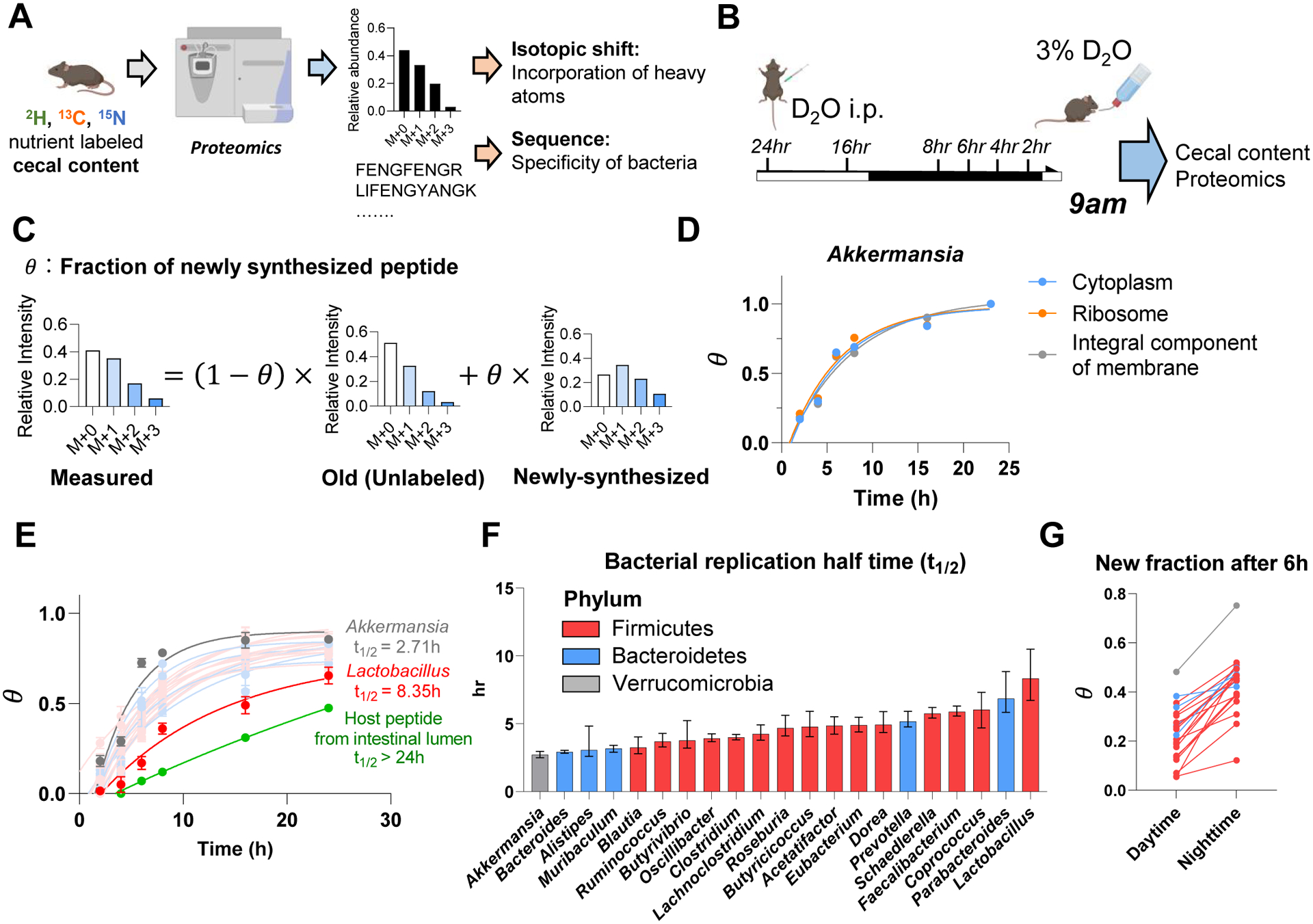
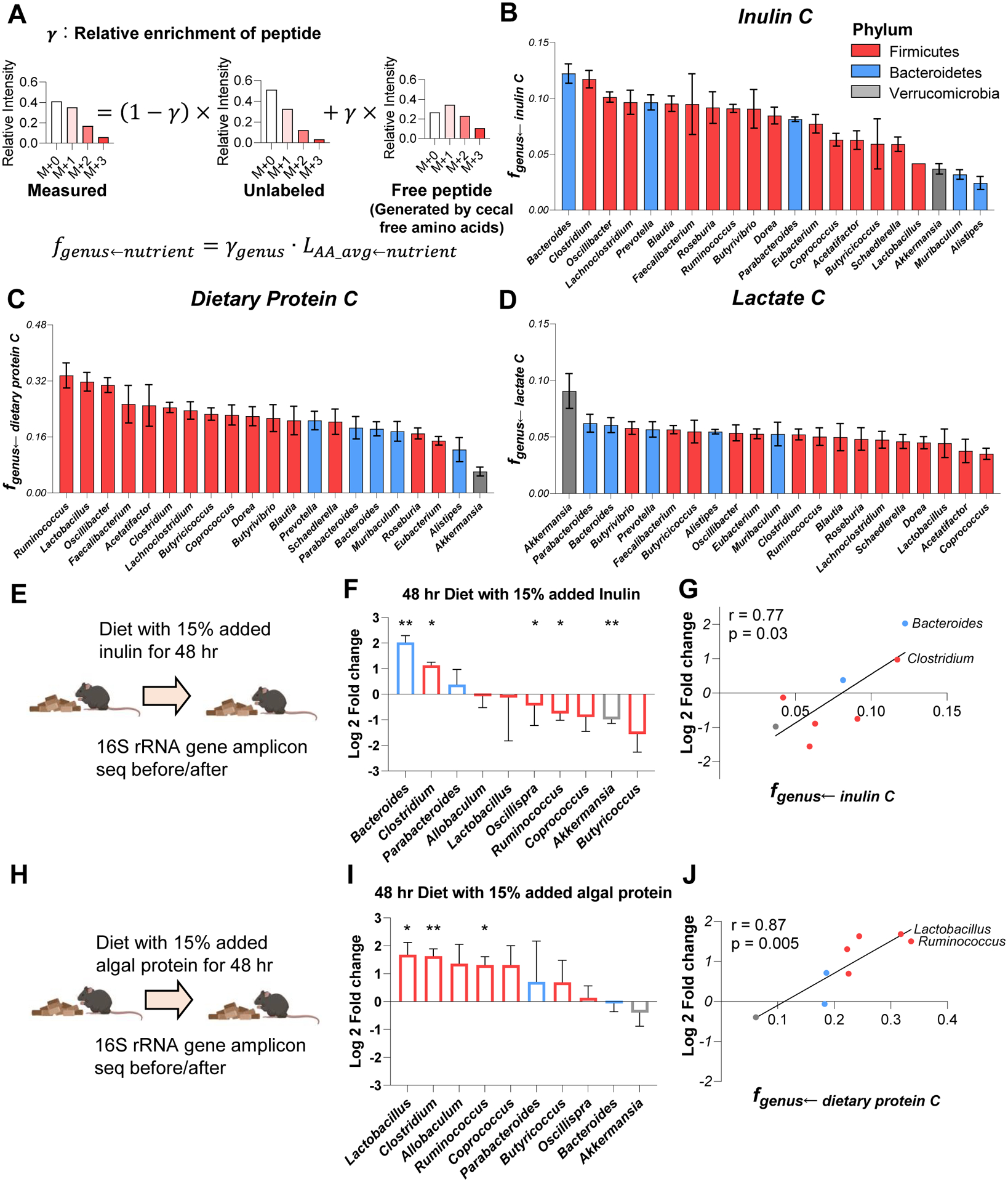
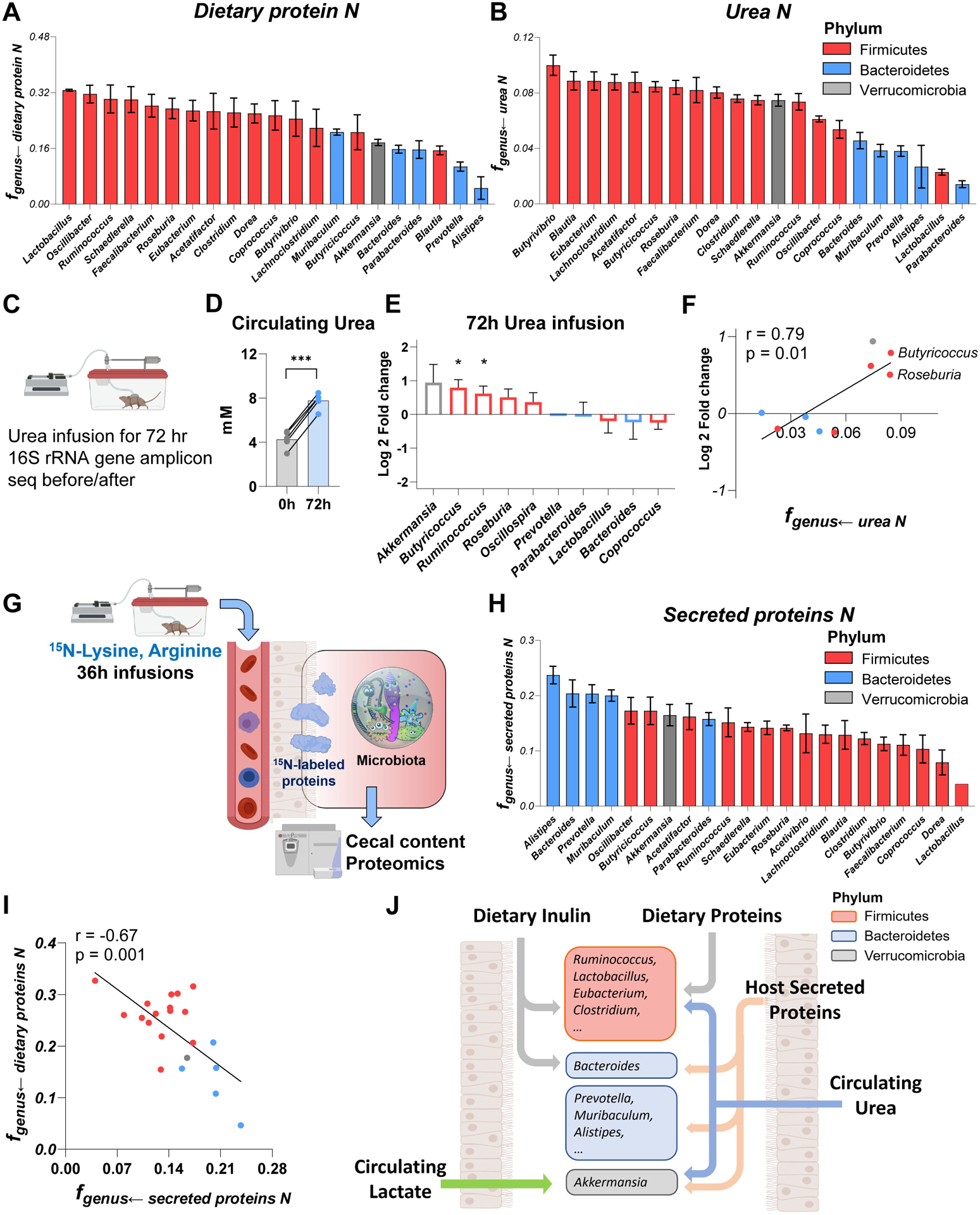
Great progress has been made in understanding gut microbiomes' products and their effects on health and disease. Less attention, however, has been given to the inputs that gut bacteria consume. Here, we quantitatively examine inputs and outputs of the mouse gut microbiome, using isotope tracing. The main input to microbial carbohydrate fermentation is dietary fiber and to branched-chain fatty acids and aromatic metabolites is dietary protein. In addition, circulating host lactate, 3-hydroxybutyrate, and urea (but not glucose or amino acids) feed the gut microbiome. To determine the nutrient preferences across bacteria, we traced into genus-specific bacterial protein sequences. We found systematic differences in nutrient use: most genera in the phylum Firmicutes prefer dietary protein, Bacteroides dietary fiber, and Akkermansia circulating host lactate. Such preferences correlate with microbiome composition changes in response to dietary modifications. Thus, diet shapes the microbiome by promoting the growth of bacteria that preferentially use the ingested nutrients.
Keywords: diet; host-microbiome interactions; isotope tracing; metabolism; metabolomics; methodology; mice; microbiome; nutrient; proteomics.
Copyright © 2022 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests J.D.R. is a member of the Rutgers Cancer Institute of New Jersey and the University of Pennsylvania Diabetes Research Center; a co-founder and stockholder in Empress Therapeutics and Serien Therapeutics; and an advisor and stockholder in Agios Pharmaceuticals, Bantam Pharmaceuticals, Colorado Research Partners, Rafael Pharmaceuticals, Barer Institute, and L.E.A.F. Pharmaceuticals. M.S.D. is a member of the scientific advisory boards of DeepBiome Therapeutics and VastBiome.
Figures
Comment in
-
Elucidating the nutrient preferences of mouse gut bacteria.Nat Rev Gastroenterol Hepatol. 2022 Nov;19(11):687. doi: 10.1038/s41575-022-00699-3. Nat Rev Gastroenterol Hepatol. 2022. PMID: 36175553 No abstract available.
References
-
- Albenberg L, Esipova TV, Judge CP, Bittinger K, Chen J, Laughlin A, Grunberg S, Baldassano RN, Lewis JD, Li H, Thom SR, Bushman FD, Vinogradov SA, Wu GD, 2014. Correlation Between Intraluminal Oxygen Gradient and Radial Partitioning of Intestinal Microbiota. Gastroenterology 147, 1055–1063.e8. 10.1053/j.gastro.2014.07.020 - DOI - PMC - PubMed
-
- Berry D, Mader E, Lee TK, Woebken D, Wang Y, Zhu D, Palatinszky M, Schintlmeister A, Schmid MC, Hanson BT, Shterzer N, Mizrahi I, Rauch I, Decker T, Bocklitz T, Popp J, Gibson CM, Fowler PW, Huang WE, Wagner M, 2015. Tracking heavy water (D2O) incorporation for identifying and sorting active microbial cells. Proc. Natl. Acad. Sci 112, E194–E203. 10.1073/pnas.1420406112 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
