

Substrate spectrum of PPM1D in the cellular response to DNA double-strand breaks
- PMID: 36060052
- PMCID: PMC9436757
- DOI: 10.1016/j.isci.2022.104892
Substrate spectrum of PPM1D in the cellular response to DNA double-strand breaks
Abstract
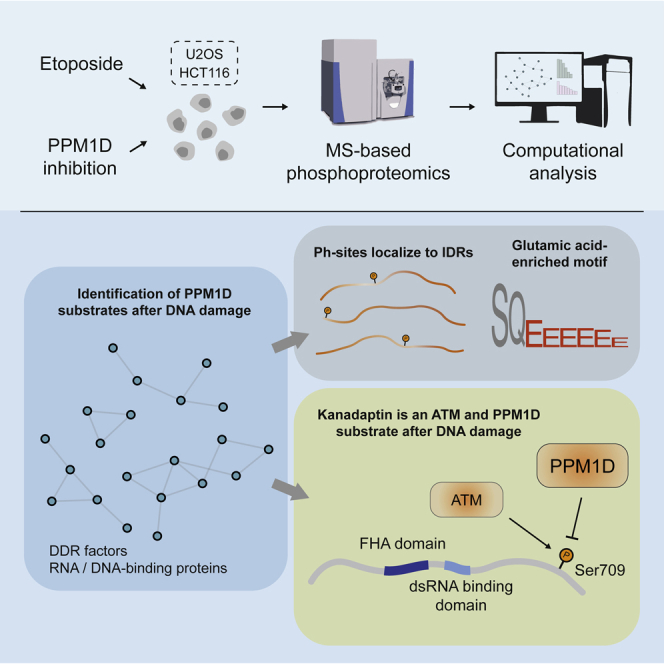
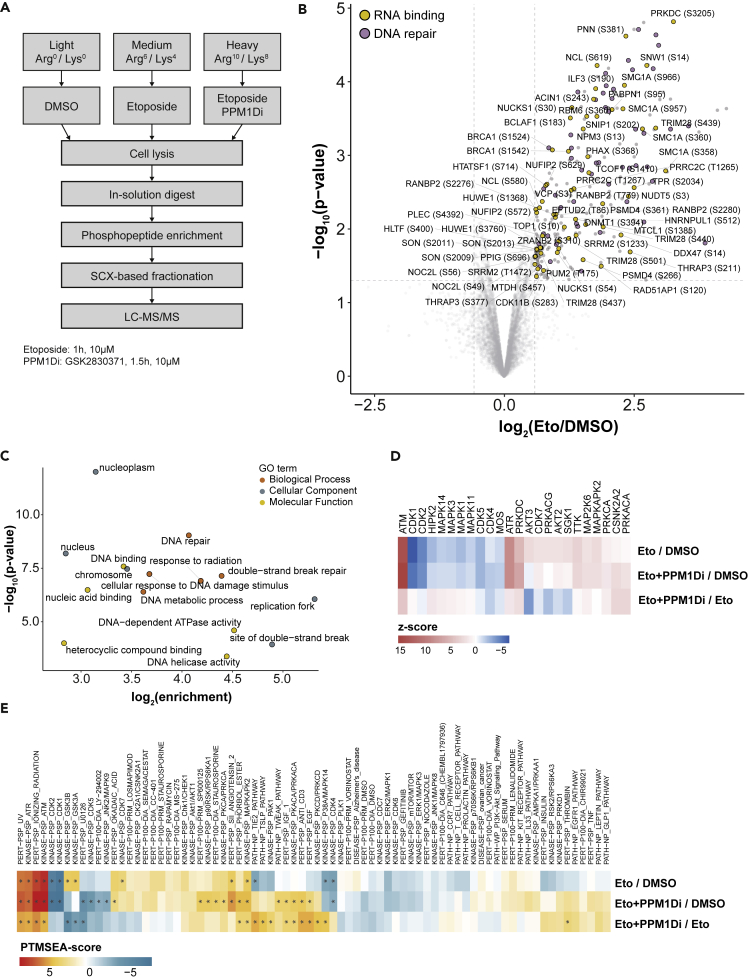
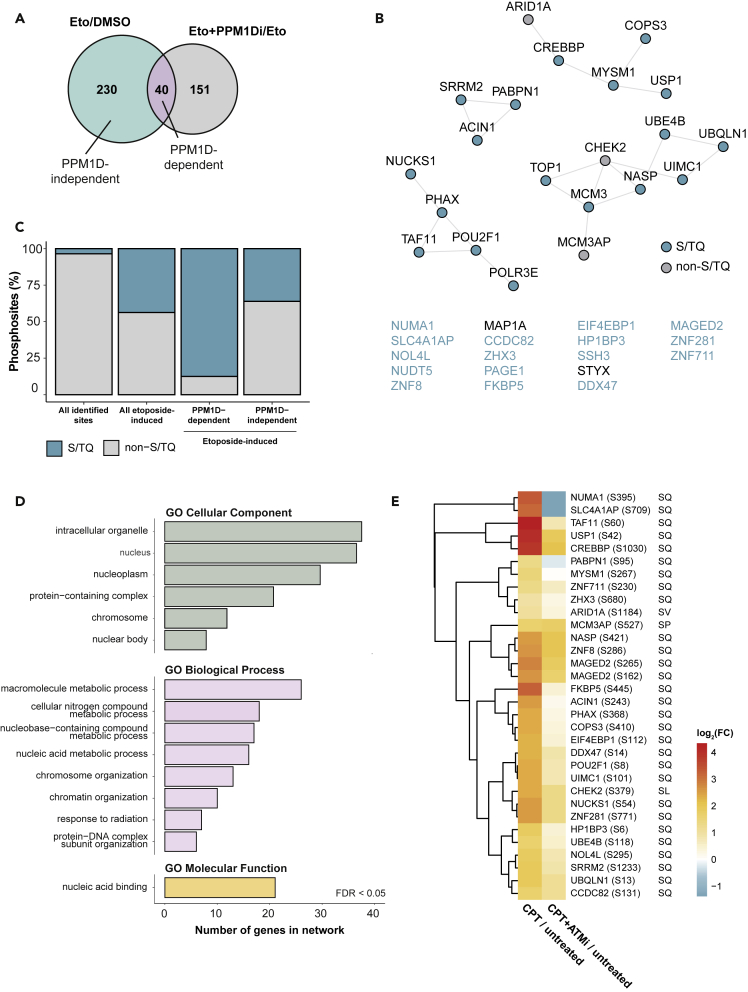
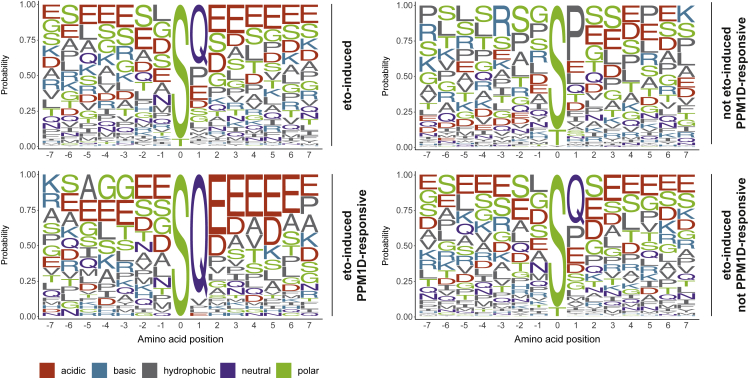
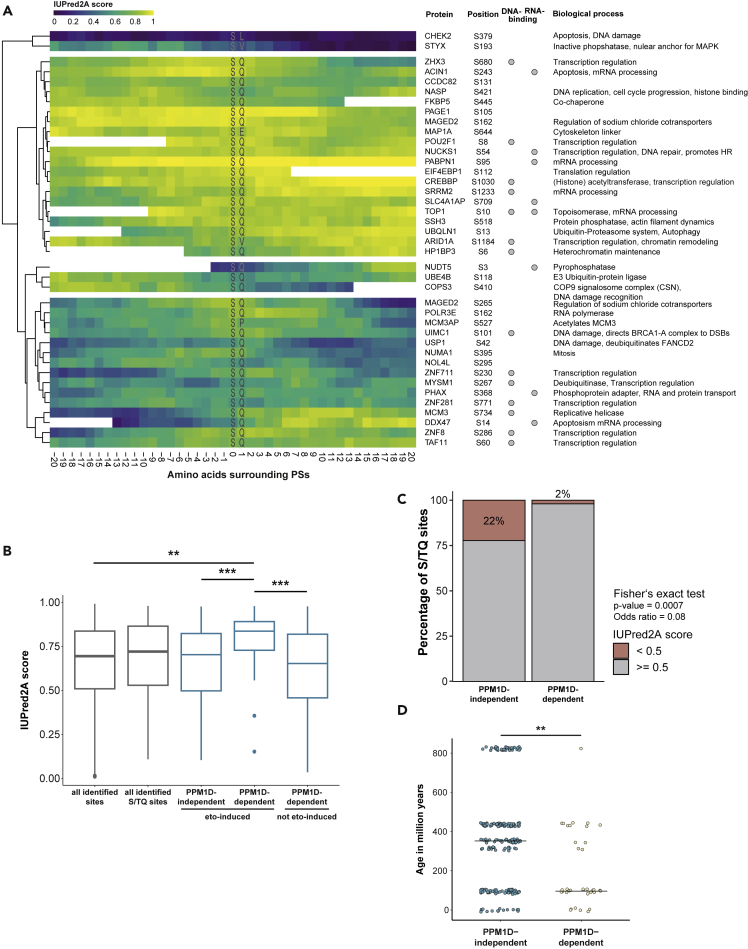
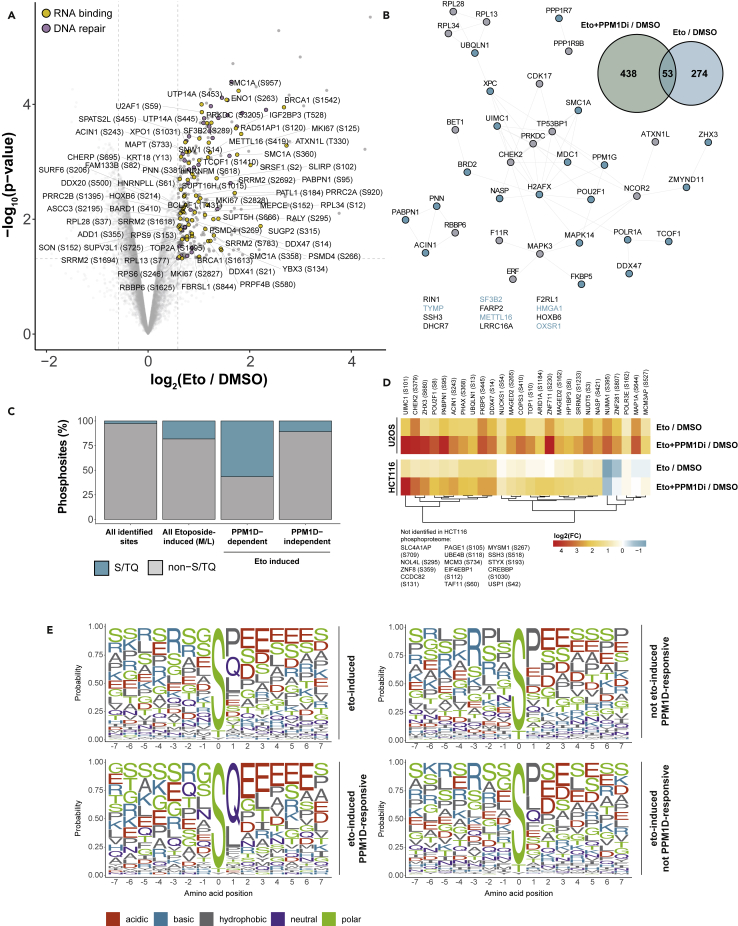
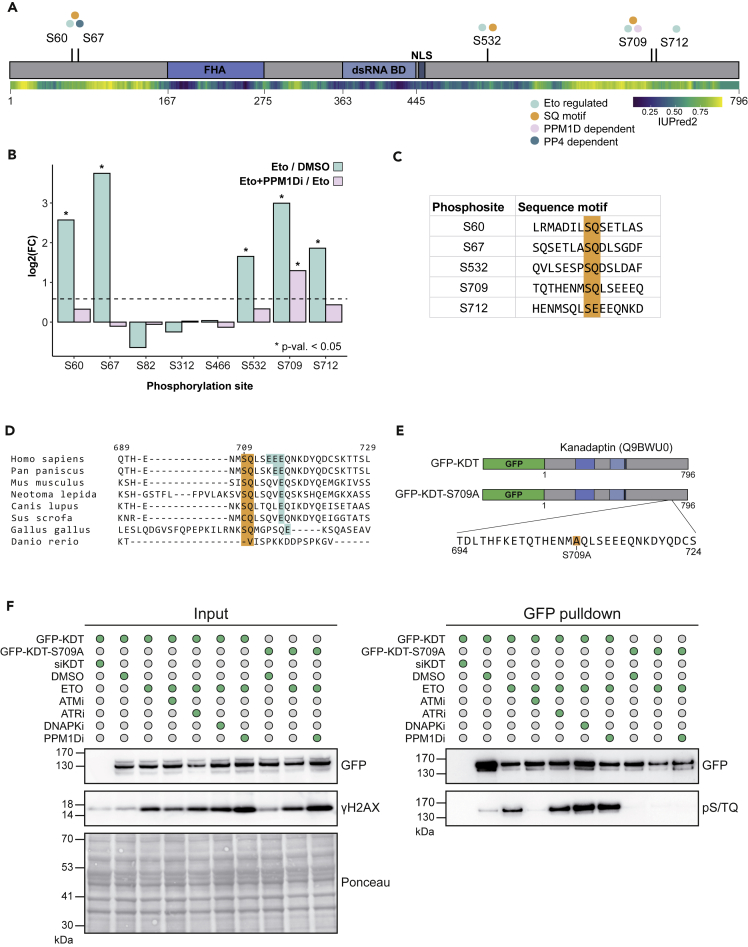
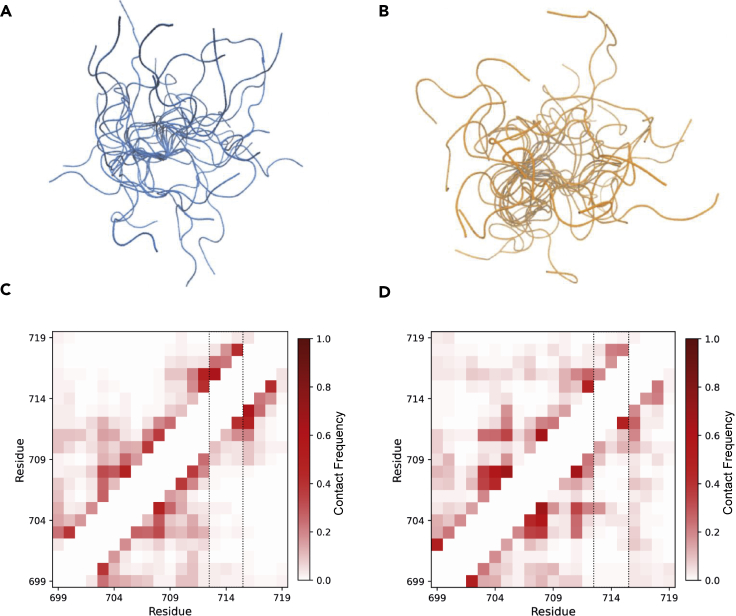
PPM1D is a p53-regulated protein phosphatase that modulates the DNA damage response (DDR) and is frequently altered in cancer. Here, we employed chemical inhibition of PPM1D and quantitative mass spectrometry-based phosphoproteomics to identify the substrates of PPM1D upon induction of DNA double-strand breaks (DSBs) by etoposide. We identified 73 putative PPM1D substrates that are involved in DNA repair, regulation of transcription, and RNA processing. One-third of DSB-induced S/TQ phosphorylation sites are dephosphorylated by PPM1D, demonstrating that PPM1D only partially counteracts ATM/ATR/DNA-PK signaling. PPM1D-targeted phosphorylation sites are found in a specific amino acid sequence motif that is characterized by glutamic acid residues, high intrinsic disorder, and poor evolutionary conservation. We identified a functionally uncharacterized protein Kanadaptin as ATM and PPM1D substrate upon DSB induction. We propose that PPM1D plays a role during the response to DSBs by regulating the phosphorylation of DNA- and RNA-binding proteins in intrinsically disordered regions.
Keywords: Biochemistry; cancer; molecular biology; proteomics.
© 2022 The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Bensimon A., Koch J.P., Francica P., Roth S.M., Riedo R., Glück A.A., Orlando E., Blaukat A., Aebersold D.M., Zimmer Y., et al. Deciphering MET-dependent modulation of global cellular responses to DNA damage by quantitative phosphoproteomics. Mol. Oncol. 2020;14:1185–1206. doi: 10.1002/1878-0261.12696. - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
