

AC5 protein encoded by squash leaf curl China virus is an RNA silencing suppressor and a virulence determinant
- PMID: 36060769
- PMCID: PMC9437540
- DOI: 10.3389/fmicb.2022.980147
AC5 protein encoded by squash leaf curl China virus is an RNA silencing suppressor and a virulence determinant
Abstract
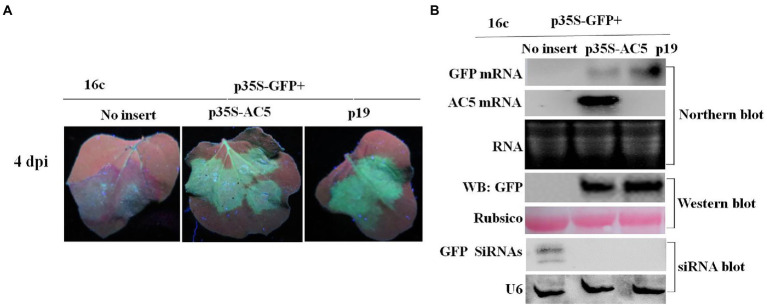
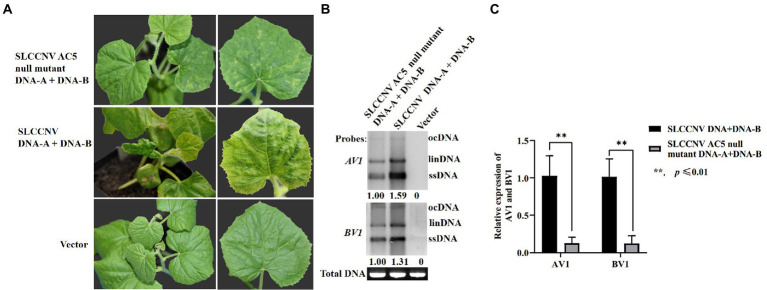
Squash leaf curl China virus (SLCCNV) is a bipartite Begomovirus. The function of the protein AC5, which is encoded by SLCCNV, is unknown. Here, we confirmed that the 172-amino acids (aa) long AC5 protein of SLCCNV could suppress single-stranded RNA but not double-stranded RNA-induced post-transcriptional gene silencing (PTGS). Furthermore, we determined that the C-terminal domain (96-172 aa) of the AC5 protein was responsible for RNA silencing suppressor (RSS) activity via deletion mutant analysis. The AC5 protein can reverse GFP silencing and inhibit systemic silencing of GFP by interfering with the systemic spread of the GFP silencing signal. The SLCCNV AC5 protein was localized to both the nucleus and cytoplasm of Nicotiana benthamiana cells. Furthermore, deletion analysis showed that the putative nuclear localization signal (NLS, 102-155 aa) was crucial for the RNA silencing suppression activity of AC5. In addition, the AC5 protein elicited a hypersensitive response and enhanced potoao virus X (PVX) RNA accumulation in infected N. benthamiana plants. Using the infectious clones of the SLCCNV and SLCCNV-AC5 null mutants, mutational analysis confirmed that knockout of the AC5 gene abolished SLCCNV-induced leaf curl symptoms, showing SLCCNV AC5 is also a virulence determinant.
Keywords: AC5 protein; RNA silencing suppressor; SLCCNV; subcellular localization; virulence determinant.
Copyright © 2022 Wu, Liu, Kang, Liu, Hong, Peng and Gu.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures






References
-
- Asigul I., Yakupjan H., Wang Y., Li H., Qian L., Han T., et al. . (2018). Cotton leaf curl Multan virus C4 protein suppresses both transcriptional and post-transcriptional gene silencing by interacting with sam synthetase. PLoS Pathog. 14:e1007282. doi: 10.1371/journal.ppat.1007282 - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources