

Interleukin-4 receptor alpha signaling regulates monocyte homeostasis
- PMID: 36063138
- PMCID: PMC9544925
- DOI: 10.1096/fj.202101672RR
Interleukin-4 receptor alpha signaling regulates monocyte homeostasis
Abstract
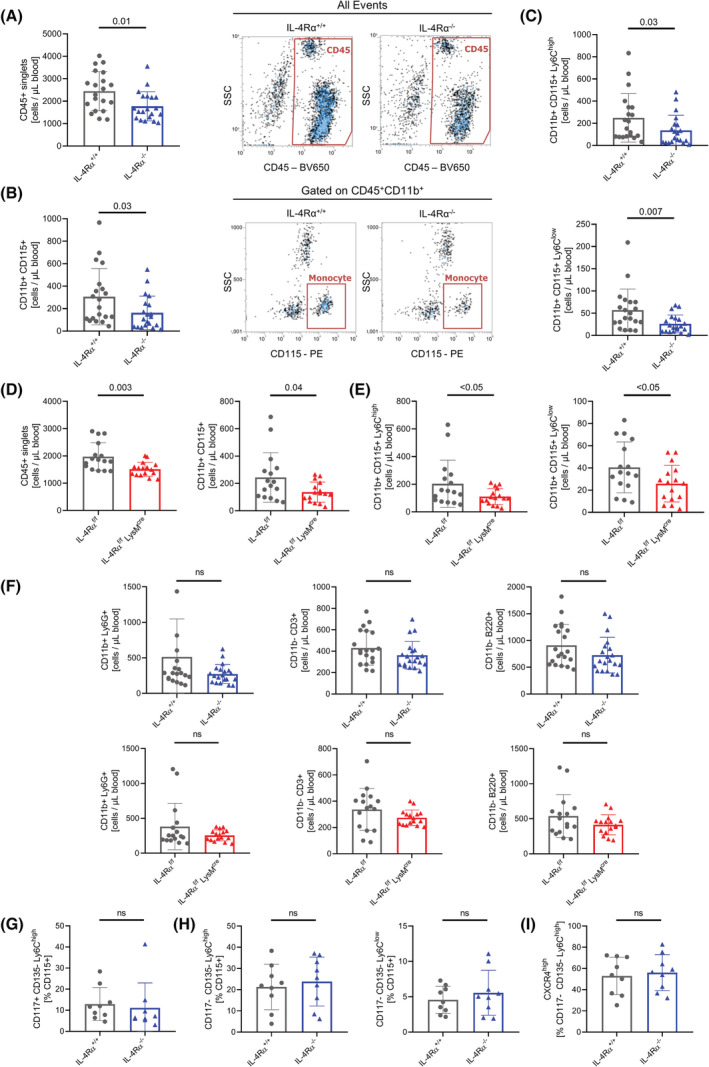
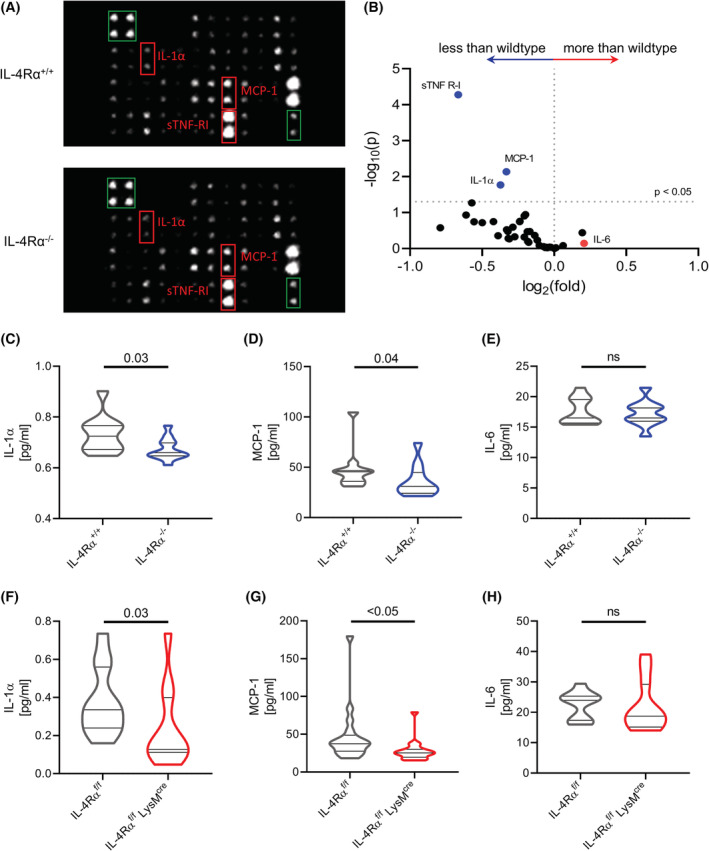
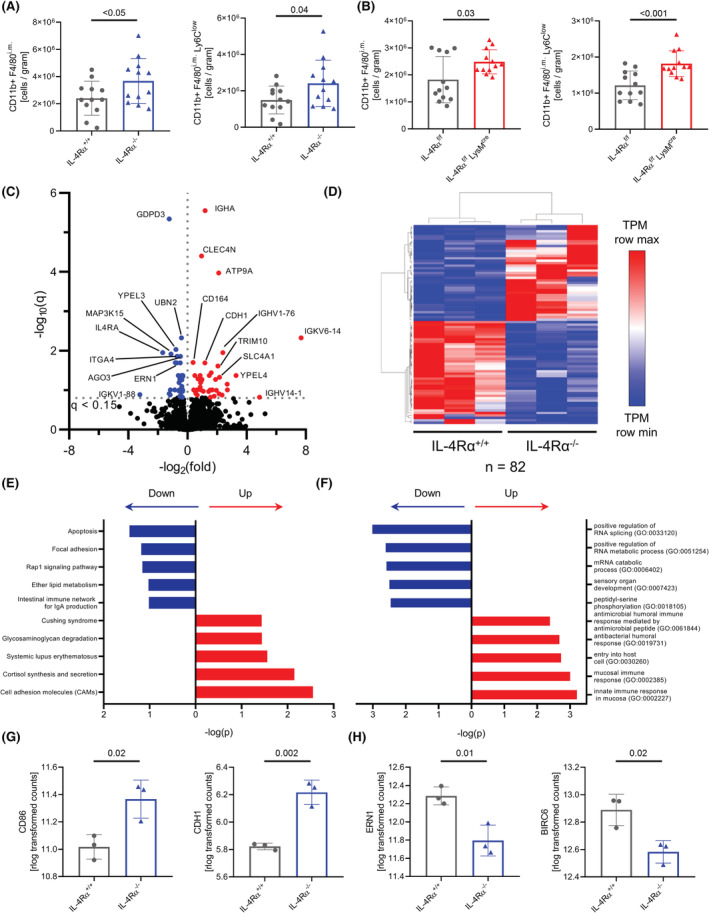
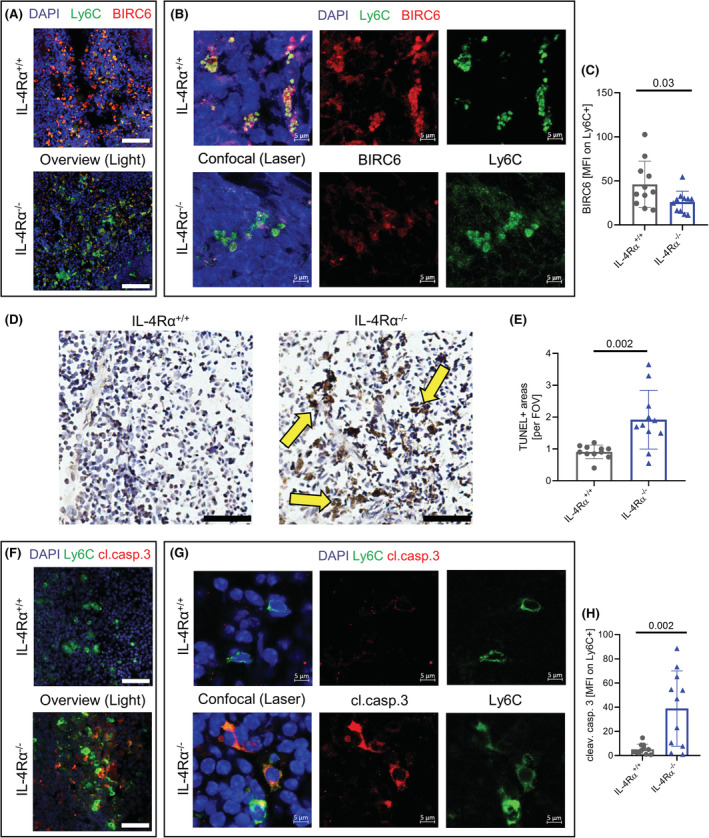
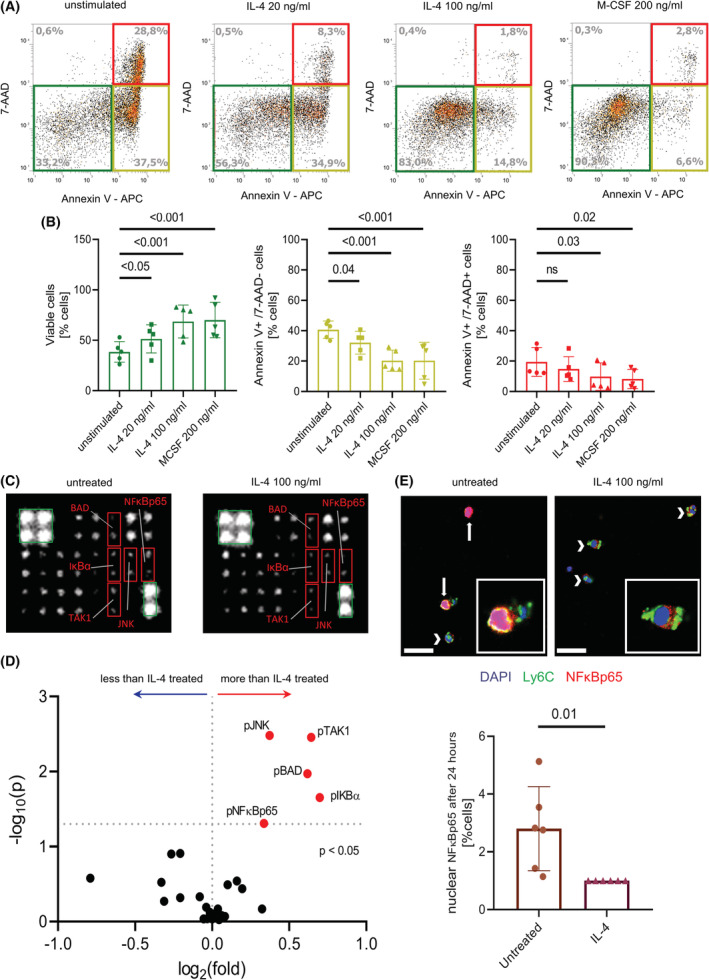
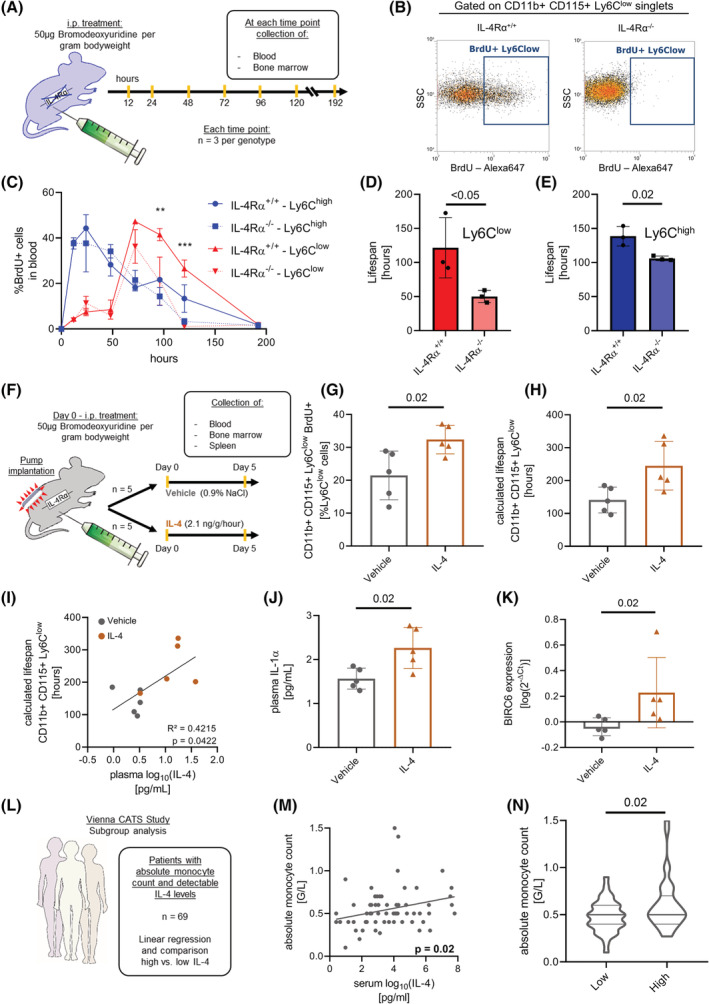
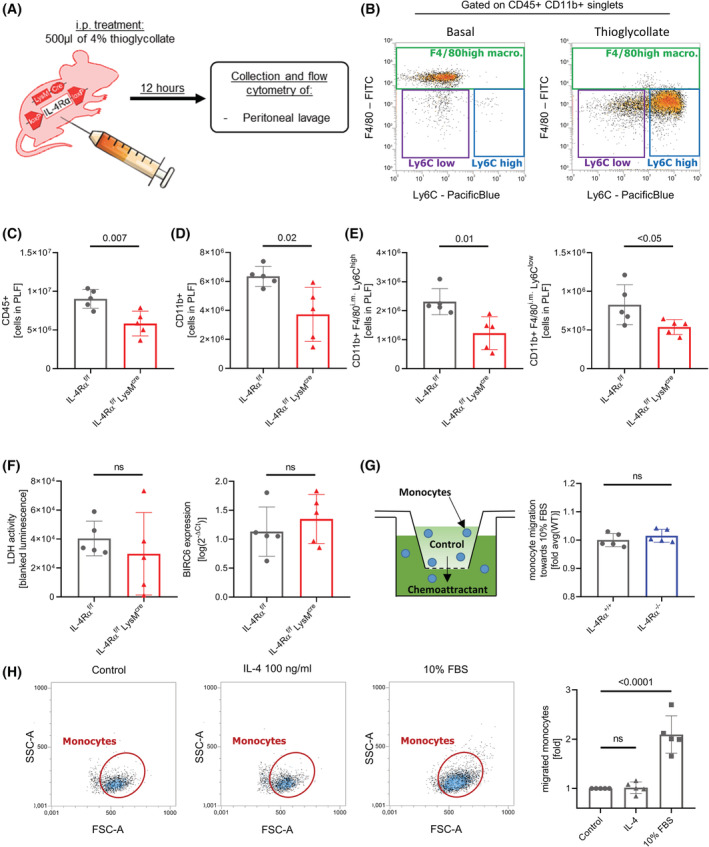
Interleukin-4 (IL-4) and its receptors (IL-4R) promote the proliferation and polarization of macrophages. However, it is unknown if IL-4R also influences monocyte homeostasis and if steady state IL-4 levels are sufficient to affect monocytes. Employing full IL-4 receptor alpha knockout mice (IL-4Rα-/- ) and mice with a myeloid-specific deletion of IL-4Rα (IL-4Rαf/f LysMcre ), we show that IL-4 acts as a homeostatic factor regulating circulating monocyte numbers. In the absence of IL-4Rα, murine monocytes in blood were reduced by 50% without altering monocytopoiesis in the bone marrow. This reduction was accompanied by a decrease in monocyte-derived inflammatory cytokines in the plasma. RNA sequencing analysis and immunohistochemical staining of splenic monocytes revealed changes in mRNA and protein levels of anti-apoptotic factors including BIRC6 in IL-4Rα-/- knockout animals. Furthermore, assessment of monocyte lifespan in vivo measuring BrdU+ cells revealed that the lifespan of circulating monocytes was reduced by 55% in IL-4Rα-/- mice, whereas subcutaneously applied IL-4 prolonged it by 75%. Treatment of human monocytes with IL-4 reduced the amount of dying monocytes in vitro. Furthermore, IL-4 stimulation reduced the phosphorylation of proteins involved in the apoptosis pathway, including the phosphorylation of the NFκBp65 protein. In a cohort of human patients, serum IL-4 levels were significantly associated with monocyte counts. In a sterile peritonitis model, reduced monocyte counts resulted in an attenuated recruitment of monocytes upon inflammatory stimulation in IL-4Rαf/f LysMcre mice without changes in overall migratory function. Thus, we identified a homeostatic role of IL-4Rα in regulating the lifespan of monocytes in vivo.
Keywords: homeostasis; immunity; innate; interleukin-4; monocytes; receptors; signal transduction.
© 2022 The Authors. The FASEB Journal published by Wiley Periodicals LLC on behalf of Federation of American Societies for Experimental Biology.
Figures
References
-
- Harris NL, Loke P. Recent advances in type‐2‐cell‐mediated immunity: insights from helminth infection. Immunity. 2017;47:1024‐1036. - PubMed
-
- Huber S, Hoffmann R, Muskens F, Voehringer D. Alternatively activated macrophages inhibit T‐cell proliferation by Stat6‐dependent expression of PD‐L2. Blood. 2010;116:3311‐3320. - PubMed
-
- Punnonen J, Yssel H, Devries J. The relative contribution of IL‐4 and IL‐13 to human IgE synthesis induced by activated CD4 or CD8 T cells. J Allergy Clin Immunol. 1997;100:792‐801. - PubMed
-
- Ranasinghe C, Roy S, Li Z, Khanna M, Jackson RJ. IL‐4 and IL‐13 receptors. In: Choi S, ed. Encyclopedia of Signaling Molecules. Springer International Publishing; 2018:2549‐2557.
-
- Bitton A, Avlas S, Reichman H, et al. A key role for IL‐13 signaling via the type 2 IL‐4 receptor in experimental atopic dermatitis. Sci Immunol. 2020;5:eaaw2938. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
