

Necroptosis is associated with Rab27-independent expulsion of extracellular vesicles containing RIPK3 and MLKL
- PMID: 36063142
- PMCID: PMC9443950
- DOI: 10.1002/jev2.12261
Necroptosis is associated with Rab27-independent expulsion of extracellular vesicles containing RIPK3 and MLKL
Abstract
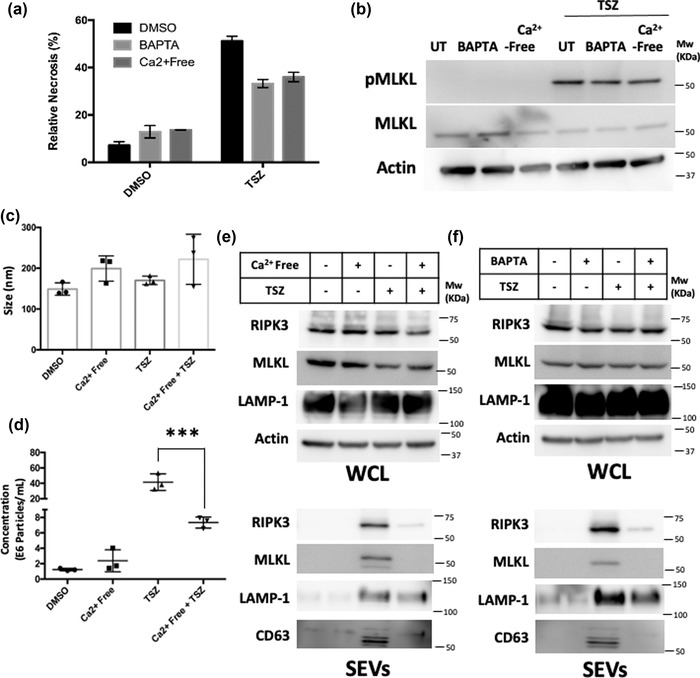
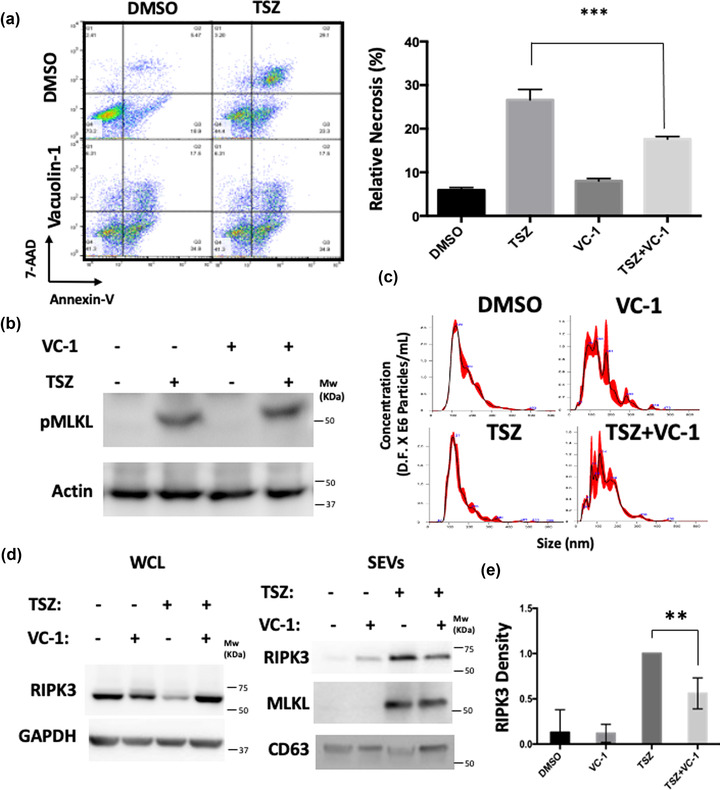
Extracellular vesicle (EV) secretion is an important mechanism used by cells to release biomolecules. A common necroptosis effector-mixed lineage kinase domain like (MLKL)-was recently found to participate in the biogenesis of small and large EVs independent of its function in necroptosis. The objective of the current study is to gain mechanistic insights into EV biogenesis during necroptosis. Assessing EV number by nanoparticle tracking analysis revealed an increased number of EVs released during necroptosis. To evaluate the nature of such vesicles, we performed a newly adapted, highly sensitive mass spectrometry-based proteomics on EVs released by healthy or necroptotic cells. Compared to EVs released by healthy cells, EVs released during necroptosis contained a markedly higher number of unique proteins. Receptor interacting protein kinase-3 (RIPK3) and MLKL were among the proteins enriched in EVs released during necroptosis. Further, mouse embryonic fibroblasts (MEFs) derived from mice deficient of Rab27a and Rab27b showed diminished basal EV release but responded to necroptosis with enhanced EV biogenesis as the wildtype MEFs. In contrast, necroptosis-associated EVs were sensitive to Ca2+ depletion or lysosomal disruption. Neither treatment affected the RIPK3-mediated MLKL phosphorylation. An unbiased screen using RIPK3 immunoprecipitation-mass spectrometry on necroptotic EVs led to the identification of Rab11b in RIPK3 immune-complexes. Our data suggests that necroptosis switches EV biogenesis from a Rab27a/b dependent mechanism to a lysosomal mediated mechanism.
Keywords: MLKL; RIPK3; SEVs; lysosomal exocytosis; necroptosis; proteomics.
© 2022 The Authors. Journal of Extracellular Vesicles published by Wiley Periodicals, LLC on behalf of the International Society for Extracellular Vesicles.
Conflict of interest statement
KAB and YG are co‐inventors on a patent that covers the detergent Azo. Other authors have no conflict of interest to declare.
Figures






References
-
- Alexander, M. , Ramstead, A. G. , Bauer, K. M. , Lee, S. ‐ H. , Runtsch, M. C. , Wallace, J. , Huffaker, T. B. , Larsen, D. K. , Tolmachova, T. , Seabra, M. C. , Round, J. L. , Ward, D. M. , & O'Connell, R. M. (2017). Rab27‐dependent exosome production inhibits chronic inflammation and enables acute responses to inflammatory stimuli. Journal of Immunology, 199, 3559–3570. 10.4049/jimmunol.1700904 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
