

A Recombinant VSV-Based Bivalent Vaccine Effectively Protects against Both SARS-CoV-2 and Influenza A Virus Infection
- PMID: 36069551
- PMCID: PMC9517730
- DOI: 10.1128/jvi.01337-22
A Recombinant VSV-Based Bivalent Vaccine Effectively Protects against Both SARS-CoV-2 and Influenza A Virus Infection
Abstract
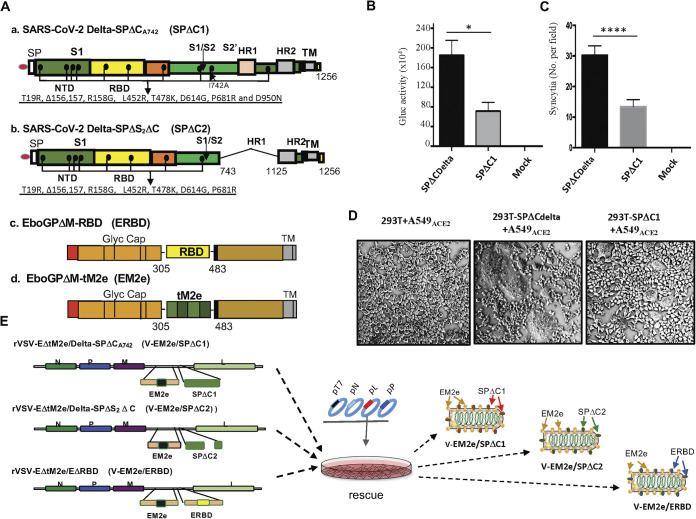
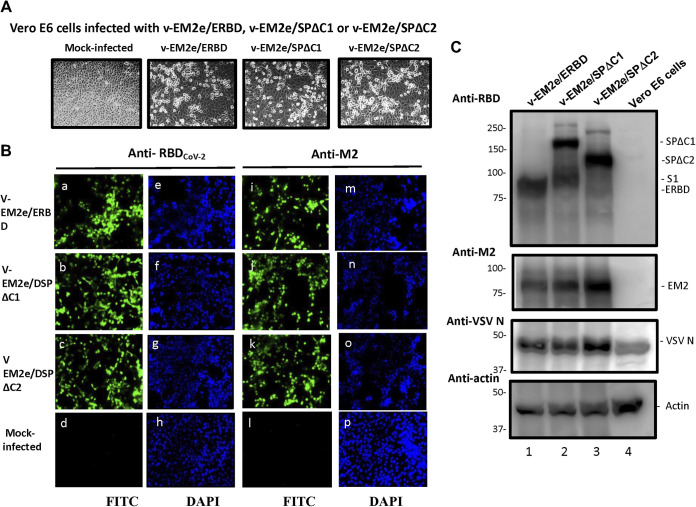
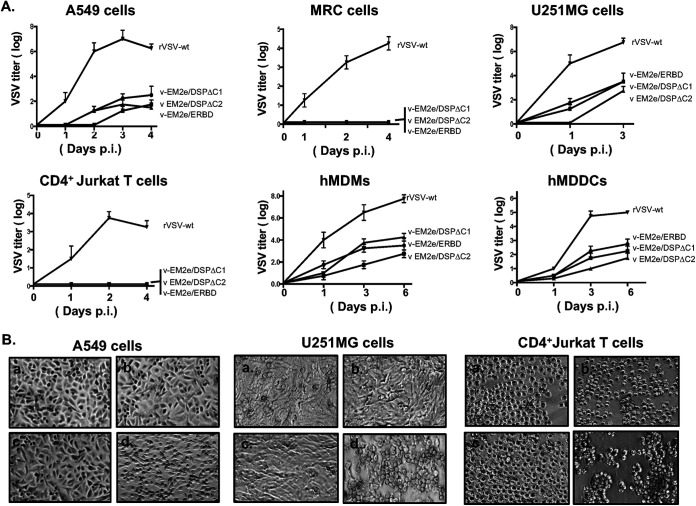
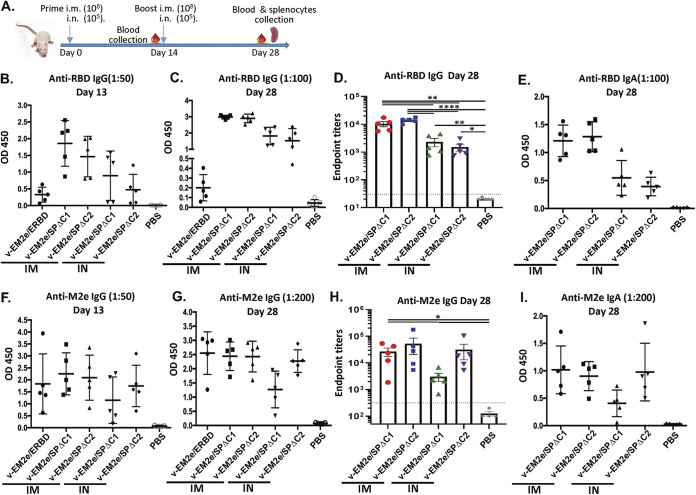
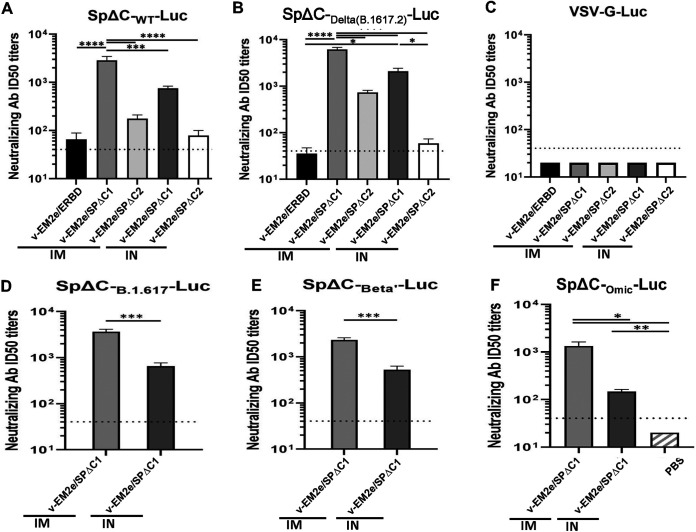
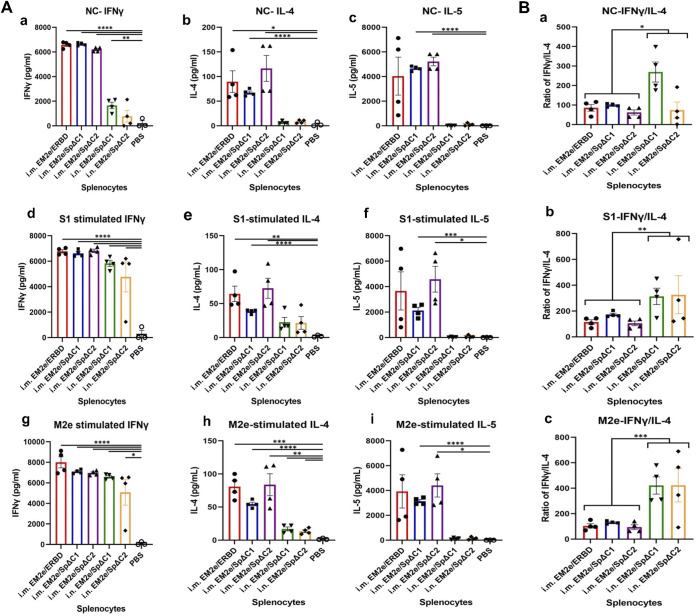
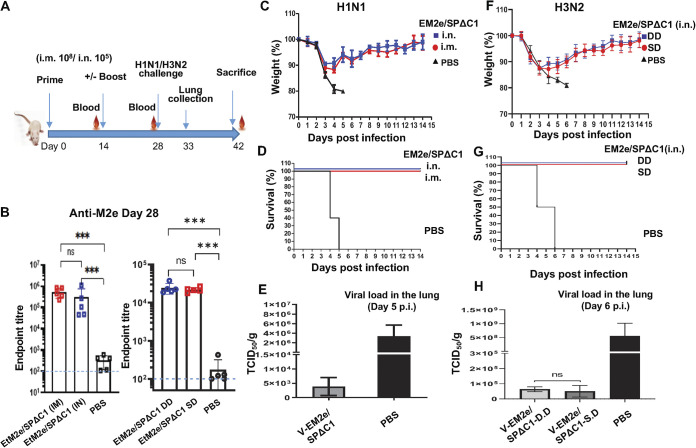
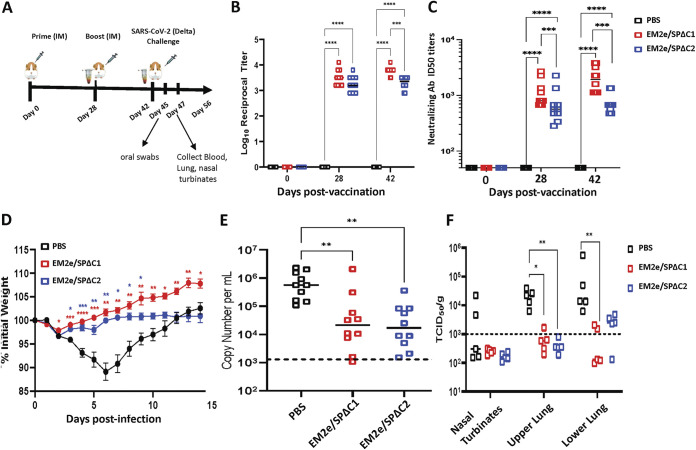
COVID-19 and influenza are both highly contagious respiratory diseases that have been serious threats to global public health. It is necessary to develop a bivalent vaccine to control these two infectious diseases simultaneously. In this study, we generated three attenuated replicating recombinant vesicular stomatitis virus (rVSV)-based vaccine candidates against both SARS-CoV-2 and influenza viruses. These rVSV-based vaccines coexpress SARS-CoV-2 Delta spike protein (SP) bearing the C-terminal 17 amino acid (aa) deletion (SPΔC) and I742A point mutation, or the SPΔC with a deletion of S2 domain, or the RBD domain, and a tandem repeat harboring four copies of the highly conserved influenza M2 ectodomain (M2e) that fused with the Ebola glycoprotein DC-targeting/activation domain. Animal immunization studies have shown that these rVSV bivalent vaccines induced efficient humoral and cellular immune responses against both SARS-CoV-2 SP and influenza M2 protein, including high levels of neutralizing antibodies against SARS-CoV-2 Delta and other variant SP-pseudovirus infections. Importantly, immunization of the rVSV bivalent vaccines effectively protected hamsters or mice against the challenges of SARS-CoV-2 Delta variant and lethal H1N1 and H3N2 influenza viruses and significantly reduced respiratory viral loads. Overall, this study provides convincing evidence for the high efficacy of this bivalent vaccine platform to be used and/or easily adapted to produce new vaccines against new or reemerging SARS-CoV-2 variants and influenza A virus infections. IMPORTANCE Given that both COVID-19 and influenza are preferably transmitted through respiratory droplets during the same seasons, it is highly advantageous to develop a bivalent vaccine that could simultaneously protect against both COVID-19 and influenza. In this study, we generated the attenuated replicating recombinant vesicular stomatitis virus (rVSV)-based vaccine candidates that target both spike protein of SARS-Cov-2 Delta variant and the conserved influenza M2 domain. Importantly, these vaccine candidates effectively protected hamsters or mice against the challenges of SARS-CoV-2 Delta variant and lethal H1N1 and H3N2 influenza viruses and significantly reduced respiratory viral loads.
Keywords: M2 protein; SARS-CoV-2 Delta variant; VSV vector; bivalent vaccine; ectodomain; influenza; neutralizing antibody; spike protein.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
