

A spatiotemporal transcriptomic network dynamically modulates stalk development in maize
- PMID: 36070002
- PMCID: PMC9674325
- DOI: 10.1111/pbi.13909
A spatiotemporal transcriptomic network dynamically modulates stalk development in maize
Abstract
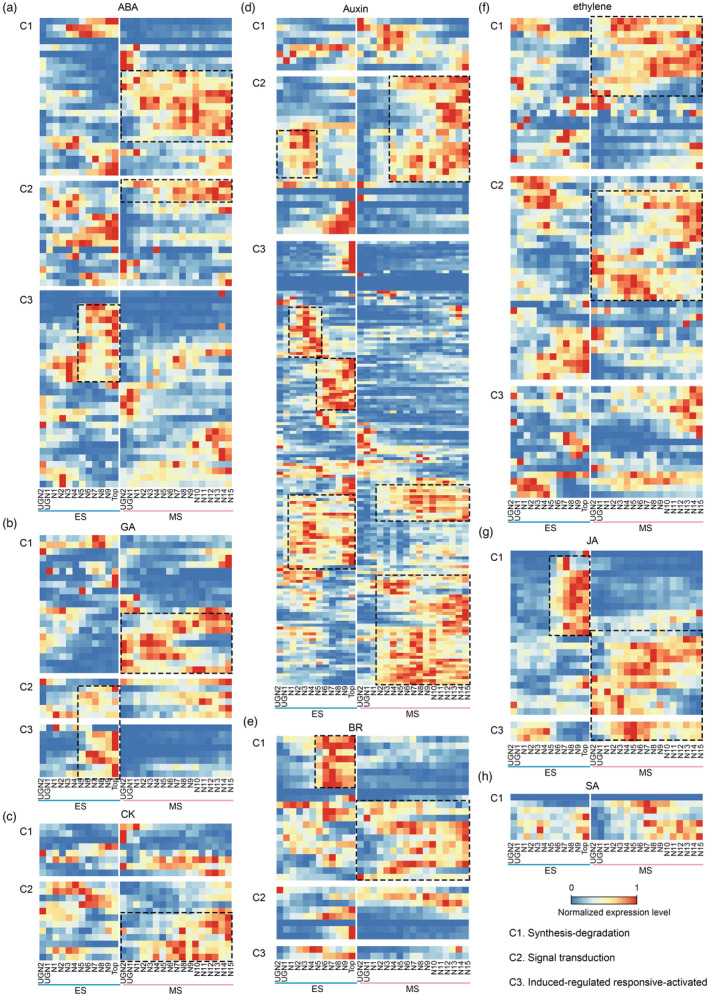
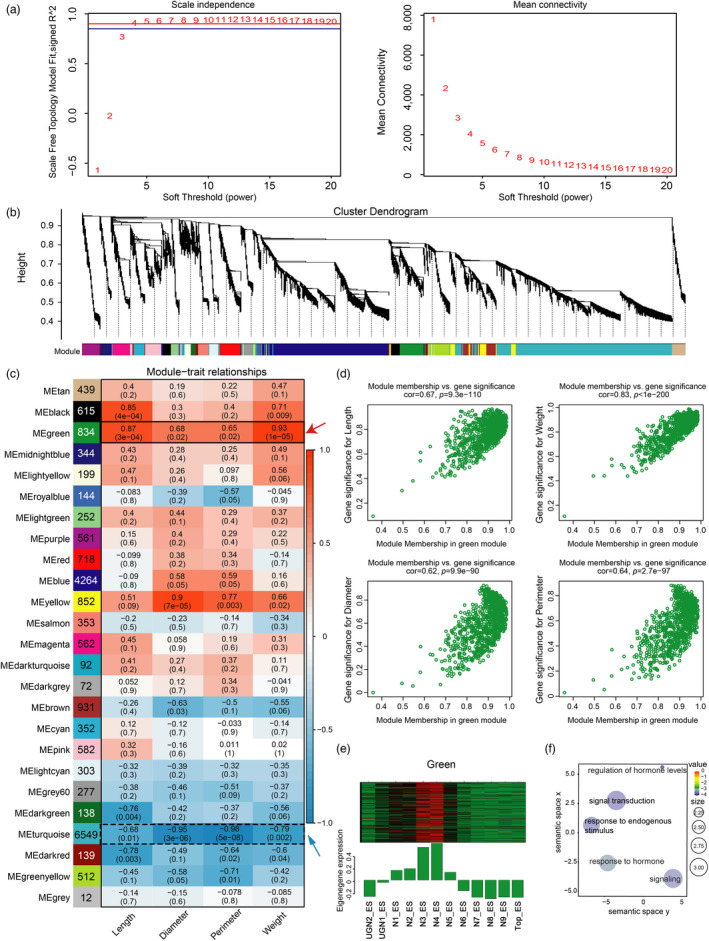
Maize (Zea mays) is an important cereal crop with suitable stalk formation which is beneficial for acquiring an ideal agronomic trait to resist lodging and higher planting density. The elongation pattern of stalks arises from the variable growth of individual internodes driven by cell division and cell expansion comprising the maize stalk. However, the spatiotemporal dynamics and regulatory network of the maize stalk development and differentiation process remain unclear. Here, we report spatiotemporally resolved transcriptomes using all internodes of the whole stalks from developing maize at the elongation and maturation stages. We identified four distinct groups corresponding to four developmental zones and nine specific clusters with diverse spatiotemporal expression patterns among individual internodes of the stalk. Through weighted gene coexpression network analysis, we constructed transcriptional regulatory networks at a fine spatiotemporal resolution and uncovered key modules and candidate genes involved in internode maintenance, elongation, and division that determine stalk length and thickness in maize. Further CRISPR/Cas9-mediated knockout validated the function of a cytochrome P450 gene, ZmD1, in the regulation of stalk length and thickness as predicted by the WGCN. Collectively, these results provide insights into the high genetic complexity of stalk development and the potentially valuable resources with ideal stalk lengths and widths for genetic improvements in maize.
Keywords: ZmD1; maize; plant height; stalk; transcriptome.
© 2022 The Authors. Plant Biotechnology Journal published by Society for Experimental Biology and The Association of Applied Biologists and John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures







References
-
- Bleecker, A.B. , Schuette, J.L. and Kende, H. (1986) Anatomical analysis of growth and developmental patterns in the internode of deepwater rice. Planta, 169, 490–497. - PubMed
-
- Cassani, E. , Bertolini, E. , Cerino Badone, F. , Landoni, M. , Gavina, D. , Sirizzotti, A. and Pilu, R. (2009) Characterization of the first dominant dwarf maize mutant carrying a single amino acid insertion in the VHYNP domain of the dwarf8 gene. Mol. Breed. 24, 375–385.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
