

Enteric glial cells favor accumulation of anti-inflammatory macrophages during the resolution of muscularis inflammation
- PMID: 36071145
- PMCID: PMC9705256
- DOI: 10.1038/s41385-022-00563-2
Enteric glial cells favor accumulation of anti-inflammatory macrophages during the resolution of muscularis inflammation
Abstract
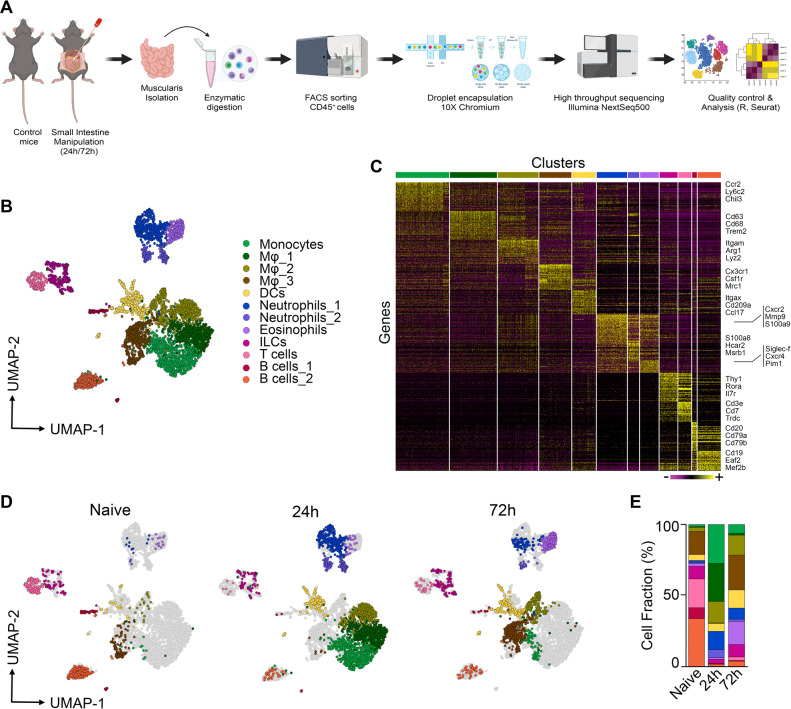
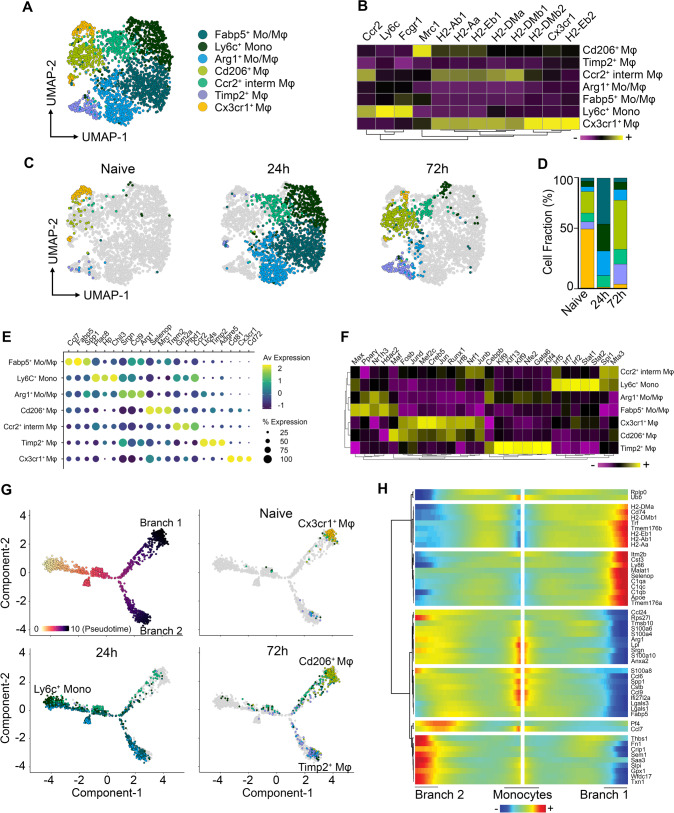
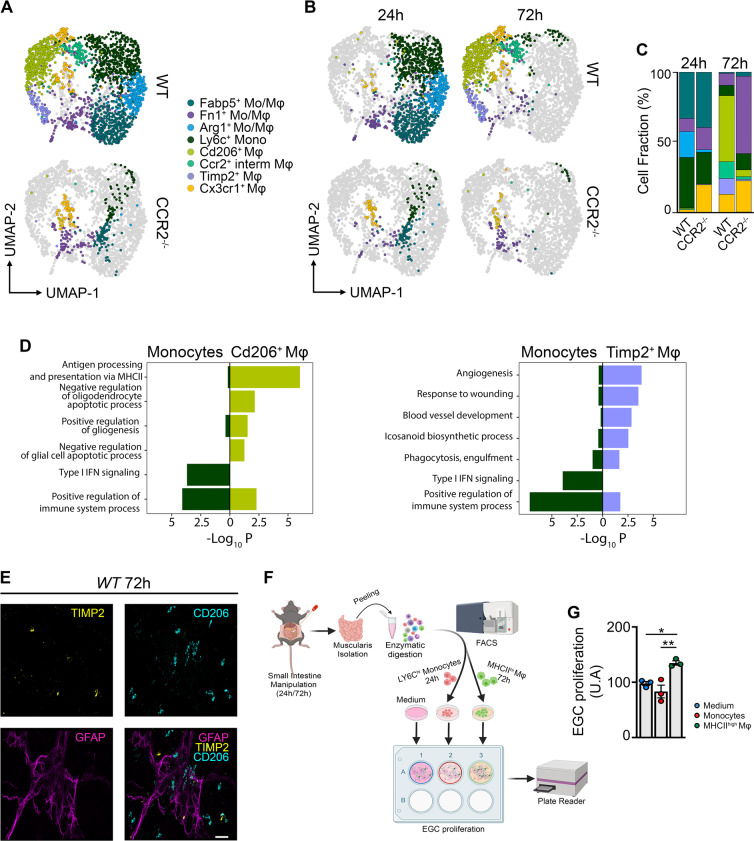
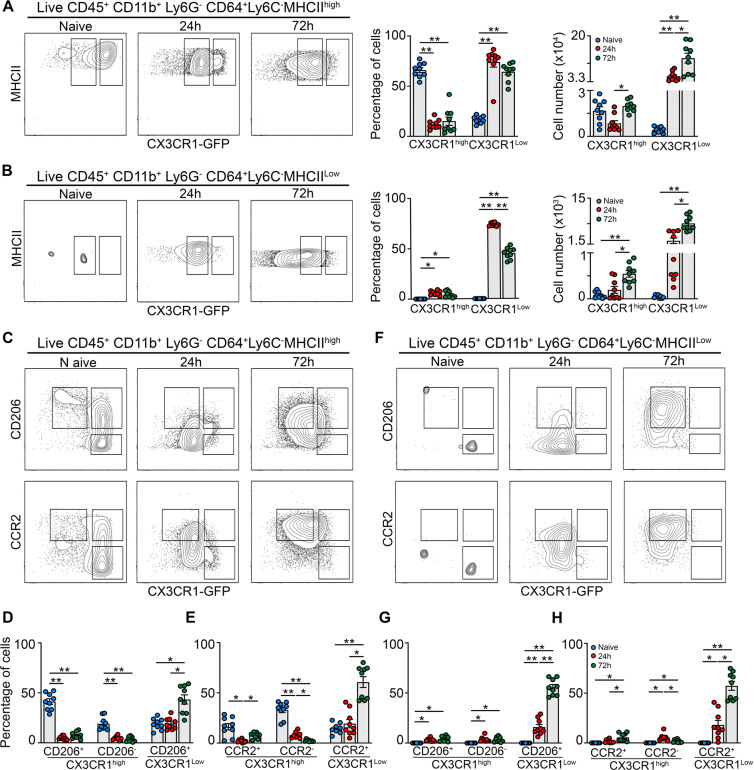
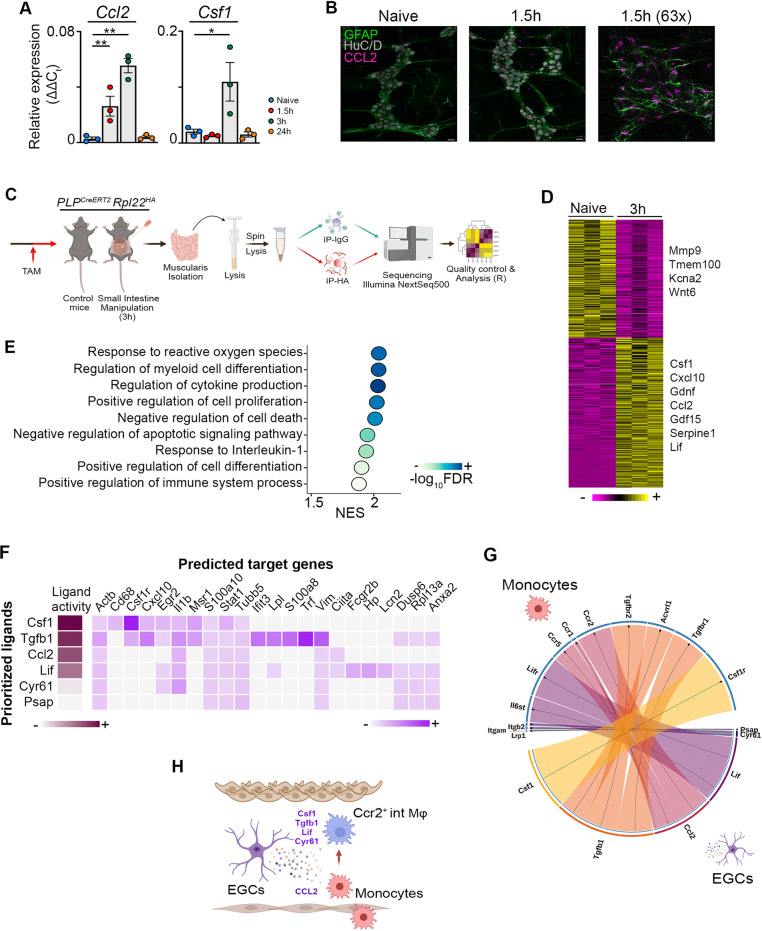
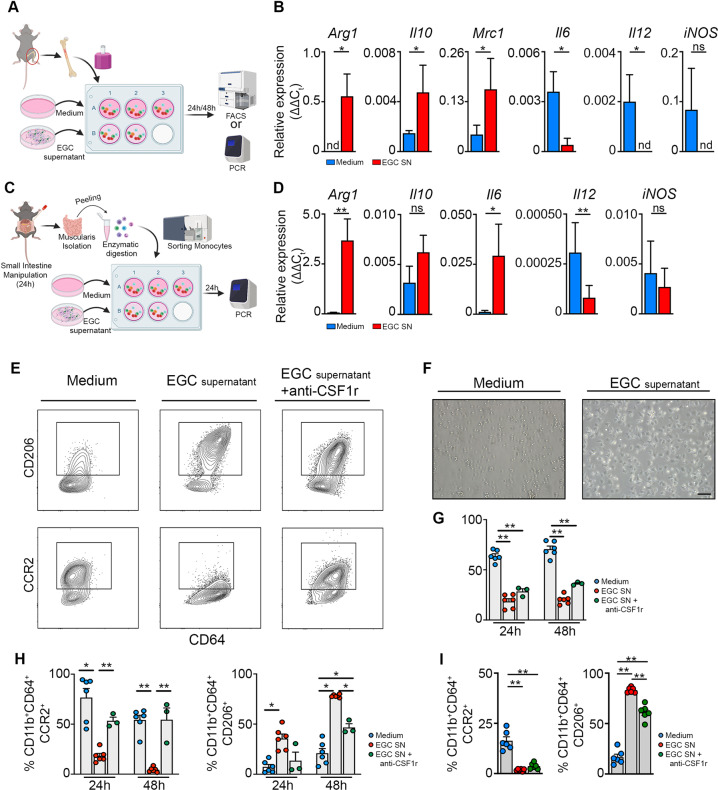
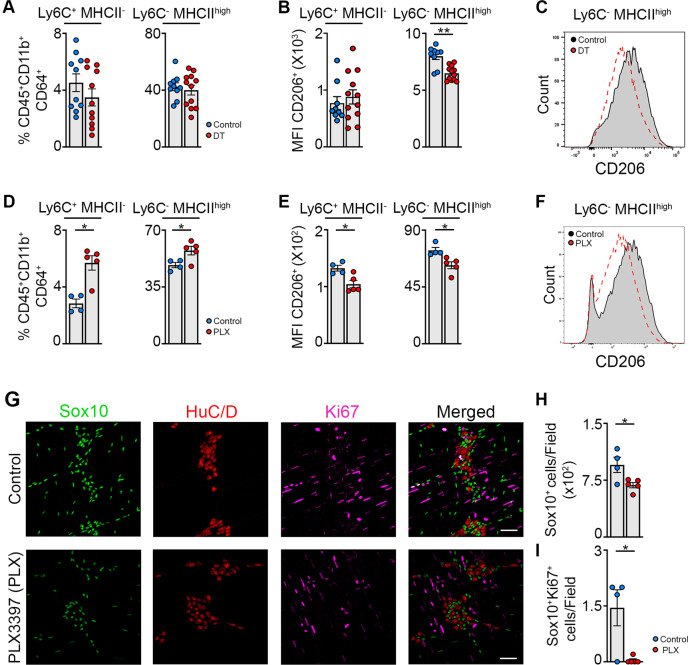
Monocyte-derived macrophages (Mφs) are crucial regulators during muscularis inflammation. However, it is unclear which micro-environmental factors are responsible for monocyte recruitment and anti-inflammatory Mφ differentiation in this paradigm. Here, we investigate Mφ heterogeneity at different stages of muscularis inflammation and determine how environmental cues can attract and activate tissue-protective Mφs. Results showed that muscularis inflammation induced marked alterations in mononuclear phagocyte populations associated with a rapid infiltration of Ly6c+ monocytes that locally acquired unique transcriptional states. Trajectory inference analysis revealed two main pro-resolving Mφ subpopulations during the resolution of muscularis inflammation, i.e. Cd206+ MhcIIhi and Timp2+ MhcIIlo Mφs. Interestingly, we found that damage to the micro-environment upon muscularis inflammation resulted in EGC activation, which in turn stimulated monocyte infiltration and the consequent differentiation in anti-inflammatory CD206+ Mφs via CCL2 and CSF1, respectively. In addition, CSF1-CSF1R signaling was shown to be essential for the differentiation of monocytes into CD206+ Mφs and EGC proliferation during muscularis inflammation. Our study provides a comprehensive insight into pro-resolving Mφ differentiation and their regulators during muscularis inflammation. We deepened our understanding in the interaction between EGCs and Mφs, thereby highlighting pro-resolving Mφ differentiation as a potential novel therapeutic strategy for the treatment of intestinal inflammation.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
