

Triblock copolymer micelle model of spherical paraspeckles
- PMID: 36072433
- PMCID: PMC9441768
- DOI: 10.3389/fmolb.2022.925058
Triblock copolymer micelle model of spherical paraspeckles
Abstract
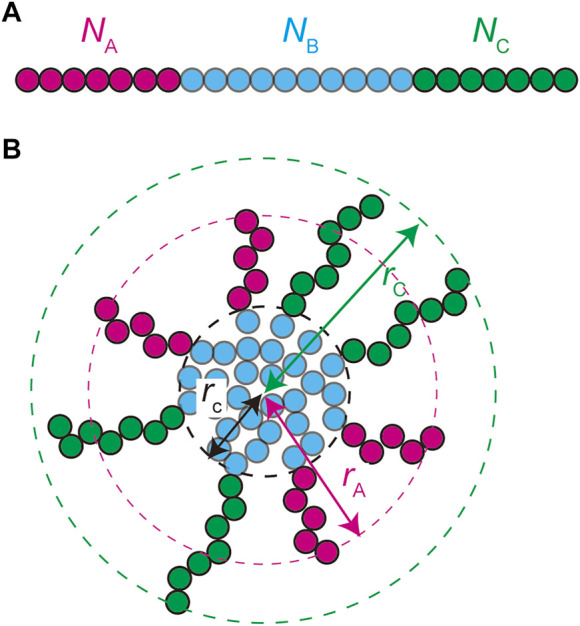
Paraspeckles are nuclear bodies scaffolded by RNP complexes of NEAT1_2 RNA transcripts and multiple RNA-binding proteins. The assembly of paraspeckles is coupled with the transcription of NEAT1_2. Paraspeckles form the core-shell structure, where the two terminal regions of NEAT1_2 RNP complexes compose the shell of the paraspeckle and the middle regions of these complexes compose the core. We here construct a theoretical model of paraspeckles by taking into account the transcription of NEAT1_2 in an extension of the theory of block copolymer micelles. This theory predicts that the core-shell structure of a paraspeckle is assembled by the association of the middle region of NEAT1_2 RNP complexes due to the multivalent interactions between RBPs bound to these regions and by the relative affinity of the terminal regions of the complexes to the nucleoplasm. The latter affinity results in the effective repulsive interactions between terminal regions of the RNA complexes and limits the number of complexes composing the paraspeckle. In the wild type, the repulsive interaction between the middle and terminal block dominates the thermal fluctuation. However, the thermal fluctuation can be significant in a mutant, where a part of the terminal regions of NEAT1_2 is deleted, and distributes the shortened terminal regions randomly between the shell and the core, consistent with our recent experiments. With the upregulated transcription, the shortened terminal regions of NEAT1_2 in a deletion mutant is localized to the core to decrease the repulsive interaction between the terminal regions, while the structure does not change with the upregulation in the wild type. The robustness of the structure of paraspeckles in the wild type results from the polymeric nature of NEAT1_2 complexes.
Keywords: NEAT1_2; architectural RNA; micellization; paraspeckle; transcription.
Copyright © 2022 Yamamoto, Yamazaki and Hirose.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures








References
-
- Alexander S. (1977). Adsorption of chain molecules with a polar head a scaling description. J. Phys. Fr. 28, 983–987. 10.1051/jphys:01977003808098300 10.1051/jphys:01977003808098300 | Google Scholar - DOI
-
- Annianson E. A. G., Wall S. N. (1974). On the kinetics of step-wise micelle association. J. Phys. Chem. 78, 1024–1030. 10.1021/j100603a016 10.1021/j100603a016 | Google Scholar - DOI
-
- Banani S. F., Lee H. O., Hyman A. A., Rosen M. K. (2017). Biomolecular condensates: Organizers of cellular biochemistry. Nat. Rev. Mol. Cell. Biol. 18, 285–298. 10.1038/nrm.2017.7 PubMed Abstract | 10.1038/nrm.2017.7 | Google Scholar - DOI - PMC - PubMed
-
- Berry J., Weber S. C., Vaidya N., Haataja M., Brangwynne C. P. (2015). RNA transcription modulates phase transition-driven nuclear body assembly. Proc. Natl. Acad. Sci. U. S. A. 112, E5237–E5245. 10.1073/pnas.1509317112 PubMed Abstract | 10.1073/pnas.1509317112 | Google Scholar - DOI - PMC - PubMed
-
- Bonetti A., Agostini F., Suzuki A. M., Hashimoto K., Pascarella G., Gimenez J., et al. (2020). RADICL-seq identifies general and cell type–specific principles of genome-wide RNA-chromatin interactions. Nat. Commun. 11, 1018. 10.1038/s41467-020-14337-6 PubMed Abstract | 10.1038/s41467-020-14337-6 | Google Scholar - DOI - PMC - PubMed
Associated data
LinkOut - more resources
Full Text Sources