

CCL2: a Chemokine Potentially Promoting Early Seeding of the Latent HIV Reservoir
- PMID: 36073812
- PMCID: PMC9600577
- DOI: 10.1128/mbio.01891-22
CCL2: a Chemokine Potentially Promoting Early Seeding of the Latent HIV Reservoir
Abstract
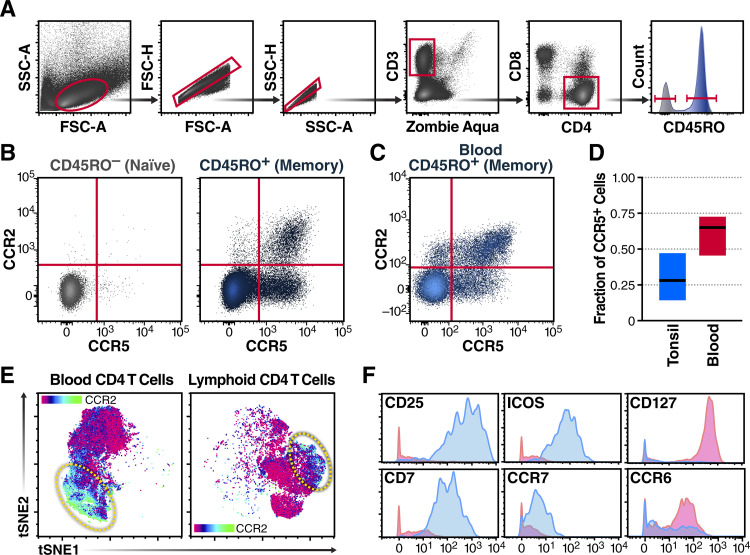
HIV infects long-lived CD4 memory T cells, establishing a latent viral reservoir that necessitates lifelong antiretroviral therapy (ART). How this reservoir is formed so quickly after infection remains unclear. We now show the innate inflammatory response to HIV infection results in CCL2 chemokine release, leading to recruitment of cells expressing the CCR2 receptor, including a subset of central memory CD4 T cells. Supporting a role for the CCL2/CCR2 axis in rapid reservoir formation, we find (i) treatment of humanized mice with anti-CCL2 antibodies during early HIV infection decreases reservoir seeding and preserves CCR2/5+ cells and (ii) CCR2/5+ cells from the blood of HIV-infected individuals on long-term ART contain significantly more integrated provirus than CCR2/5-negative memory or naive cells. Together, these studies support a model where the host's innate inflammatory response to HIV infection, including CCL2 production, leads to the recruitment of CCR2/5+ central memory CD4 T cells to zones of virus-associated inflammation, likely contributing to rapid formation of the latent HIV reservoir. IMPORTANCE There are currently over 35 million people living with HIV worldwide, and we still have no vaccine or scalable cure. One of the difficulties with HIV is its ability to rapidly establish a viral reservoir in lymphoid tissues that allows it to elude antivirals and the immune system. Thus, it is important to understand how HIV accomplishes this so we can develop preventive strategies. Our current results show that an early inflammatory response to HIV infection includes production of the chemokine CCL2, which recruits a unique subset of CCR2/5+ CD4+ T cells that become infected and form a significant reservoir for latent infection. Furthermore, we show that blockade of CCL2 in humanized mice significantly reduces persistent HIV infection. This information is relevant to the development of therapeutics to prevent and/or treat chronic HIV infections.
Keywords: CCL2; CRISPR; CyTOF; human immunodeficiency virus; latency; reservoir.
Conflict of interest statement
The authors declare no conflict of interest.
Figures





Similar articles
-
Prescription of Controlled Substances: Benefits and Risks.2025 Jul 6. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. 2025 Jul 6. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. PMID: 30726003 Free Books & Documents.
-
The role of Nef in the long-term persistence of the replication-competent HIV reservoir in South African women.J Virol. 2025 Jul 22;99(7):e0021725. doi: 10.1128/jvi.00217-25. Epub 2025 Jun 24. J Virol. 2025. PMID: 40552830 Free PMC article.
-
Establishment and Reversal of HIV-1 Latency in Naive and Central Memory CD4+ T Cells In Vitro.J Virol. 2016 Aug 26;90(18):8059-73. doi: 10.1128/JVI.00553-16. Print 2016 Sep 15. J Virol. 2016. PMID: 27356901 Free PMC article.
-
Structured treatment interruptions (STI) in chronic unsuppressed HIV infection in adults.Cochrane Database Syst Rev. 2006 Jul 19;2006(3):CD006148. doi: 10.1002/14651858.CD006148. Cochrane Database Syst Rev. 2006. PMID: 16856117 Free PMC article.
-
Antiretroviral therapy (ART) for treating HIV infection in ART-eligible pregnant women.Cochrane Database Syst Rev. 2010 Mar 17;2010(3):CD008440. doi: 10.1002/14651858.CD008440. Cochrane Database Syst Rev. 2010. PMID: 20238370 Free PMC article.
Cited by
-
Activation-neutral gene editing of tonsillar CD4 T cells for functional studies in human ex vivo tonsil cultures.Cell Rep Methods. 2024 Jan 22;4(1):100685. doi: 10.1016/j.crmeth.2023.100685. Epub 2024 Jan 10. Cell Rep Methods. 2024. PMID: 38211593 Free PMC article.
-
Macrophage-Derived Factors with the Potential to Contribute to Pathogenicity of HIV-1 and HIV-2: Role of CCL-2/MCP-1.Viruses. 2023 Oct 27;15(11):2160. doi: 10.3390/v15112160. Viruses. 2023. PMID: 38005838 Free PMC article.
-
Differential Impact of IL-32 Isoforms on the Functions of Coronary Artery Endothelial Cells: A Potential Link with Arterial Stiffness and Atherosclerosis.Viruses. 2023 Mar 8;15(3):700. doi: 10.3390/v15030700. Viruses. 2023. PMID: 36992409 Free PMC article.
-
Antiretroviral drug therapy does not reduce neuroinflammation in an HIV-1 infection brain organoid model.J Neuroinflammation. 2025 Mar 5;22(1):66. doi: 10.1186/s12974-025-03375-w. J Neuroinflammation. 2025. PMID: 40045391 Free PMC article.
-
The Humanized Mouse Model: What Added Value Does It Offer for HIV Research?Pathogens. 2023 Apr 17;12(4):608. doi: 10.3390/pathogens12040608. Pathogens. 2023. PMID: 37111494 Free PMC article. Review.
References
-
- Whitney JB, Hill AL, Sanisetty S, Penaloza-MacMaster P, Liu J, Shetty M, Parenteau L, Cabral C, Shields J, Blackmore S, Smith JY, Brinkman AL, Peter LE, Mathew SI, Smith KM, Borducchi EN, Rosenbloom DI, Lewis MG, Hattersley J, Li B, Hesselgesser J, Geleziunas R, Robb ML, Kim JH, Michael NL, Barouch DH. 2014. Rapid seeding of the viral reservoir prior to SIV viraemia in rhesus monkeys. Nature 512:74–77. doi:10.1038/nature13594. - DOI - PMC - PubMed
-
- Colby DJ, Trautmann L, Pinyakorn S, Leyre L, Pagliuzza A, Kroon E, Rolland M, Takata H, Buranapraditkun S, Intasan J, Chomchey N, Muir R, Haddad EK, Tovanabutra S, Ubolyam S, Bolton DL, Fullmer BA, Gorelick RJ, Fox L, Crowell TA, Trichavaroj R, O’Connell R, Chomont N, Kim JH, Michael NL, Robb ML, Phanuphak N, Ananworanich J, RV411 study group . 2018. Rapid HIV RNA rebound after antiretroviral treatment interruption in persons durably suppressed in Fiebig I acute HIV infection. Nat Med 24:923–926. doi:10.1038/s41591-018-0026-6. - DOI - PMC - PubMed
-
- Henrich TJ, Hatano H, Bacon O, Hogan LE, Rutishauser R, Hill A, Kearney MF, Anderson EM, Buchbinder SP, Cohen SE, Abdel-Mohsen M, Pohlmeyer CW, Fromentin R, Hoh R, Liu AY, McCune JM, Spindler J, Metcalf-Pate K, Hobbs KS, Thanh C, Gibson EA, Kuritzkes DR, Siliciano RF, Price RW, Richman DD, Chomont N, Siliciano JD, Mellors JW, Yukl SA, Blankson JN, Liegler T, Deeks SG. 2017. HIV-1 persistence following extremely early initiation of antiretroviral therapy (ART) during acute HIV-1 infection: an observational study. PLoS Med 14:e1002417. doi:10.1371/journal.pmed.1002417. - DOI - PMC - PubMed
-
- Abrahams MR, Joseph SB, Garrett N, Tyers L, Moeser M, Archin N, Council OD, Matten D, Zhou S, Doolabh D, Anthony C, Goonetilleke N, Karim SA, Margolis DM, Pond SK, Williamson C, Swanstrom R. 2019. The replication-competent HIV-1 latent reservoir is primarily established near the time of therapy initiation. Sci Transl Med 11:eaaw5589. doi:10.1126/scitranslmed.aaw5589. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
