

Mechanisms of ADAMTS13 regulation
- PMID: 36074019
- PMCID: PMC9826392
- DOI: 10.1111/jth.15873
Mechanisms of ADAMTS13 regulation
Abstract
Recombinant ADAMTS13 is currently undergoing clinical trials as a treatment for hereditary thrombotic thrombocytopenic purpura, a lethal microvascular condition resulting from ADAMTS13 deficiency. Preclinical studies have also demonstrated its efficacy in treating arterial thrombosis and inflammation without causing bleeding, suggesting that recombinant ADAMTS13 may have broad applicability as an antithrombotic agent. Despite this progress, we currently do not understand the mechanisms that regulate ADAMTS13 activity in vivo. ADAMTS13 evades canonical means of protease regulation because it is secreted as an active enzyme and has a long half-life in circulation, suggesting that it is not inhibited by natural protease inhibitors. Although shear can spatially and temporally activate von Willebrand factor to capture circulating platelets, it is also required for cleavage by ADAMTS13. Therefore, spatial and temporal regulation of ADAMTS13 activity may be required to stabilize von Willebrand factor-platelet strings at sites of vascular injury. This review outlines potential mechanisms that regulate ADAMTS13 in vivo including shear-dependency, local inactivation, and biochemical and structural regulation of substrate binding. Recently published structural data of ADAMTS13 is discussed, which may help to generate novel hypotheses for future research.
Keywords: ADAMTS13; VWF; antithrombotic; protease regulation; thrombosis.
© 2022 The Authors. Journal of Thrombosis and Haemostasis published by Wiley Periodicals LLC on behalf of International Society on Thrombosis and Haemostasis.
Conflict of interest statement
All of the authors declare that there are no conflicts of interest.
Figures
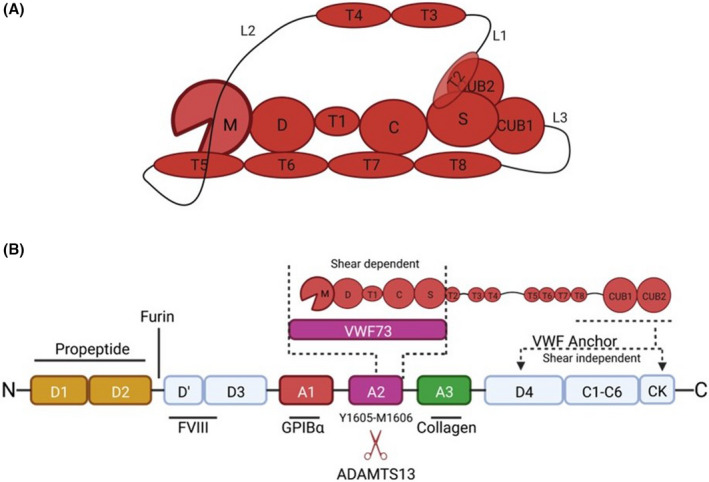
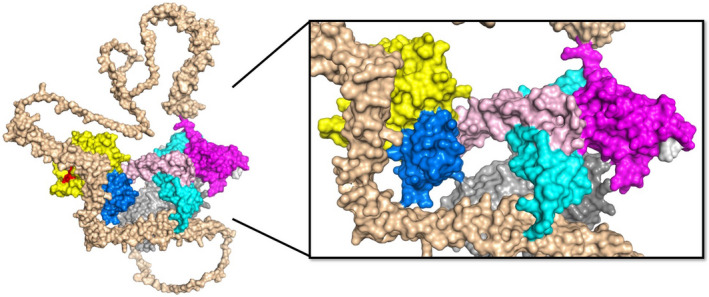
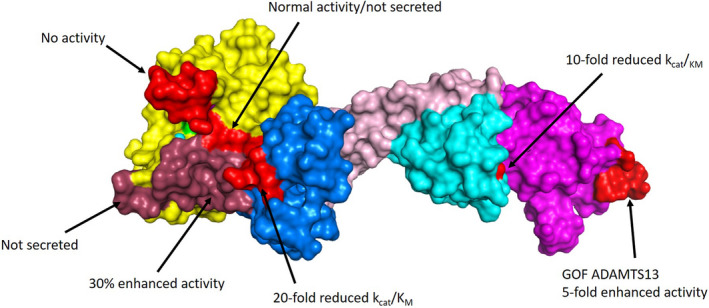
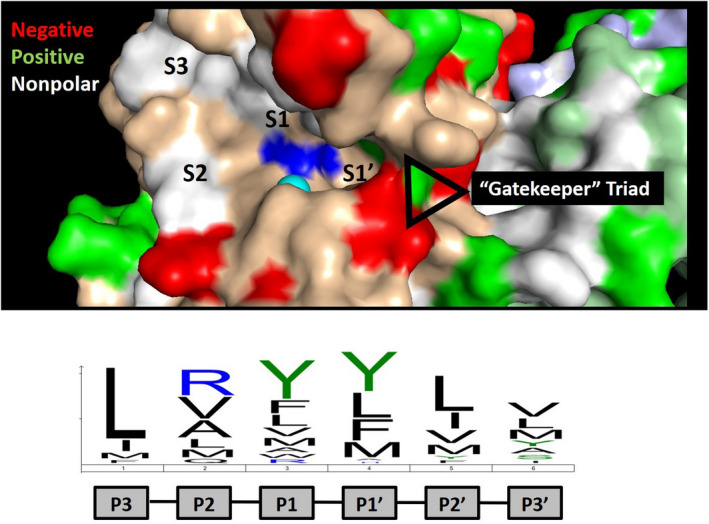
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
