

The iron-sulfur cluster assembly (ISC) protein Iba57 executes a tetrahydrofolate-independent function in mitochondrial [4Fe-4S] protein maturation
- PMID: 36075292
- PMCID: PMC9551070
- DOI: 10.1016/j.jbc.2022.102465
The iron-sulfur cluster assembly (ISC) protein Iba57 executes a tetrahydrofolate-independent function in mitochondrial [4Fe-4S] protein maturation
Abstract
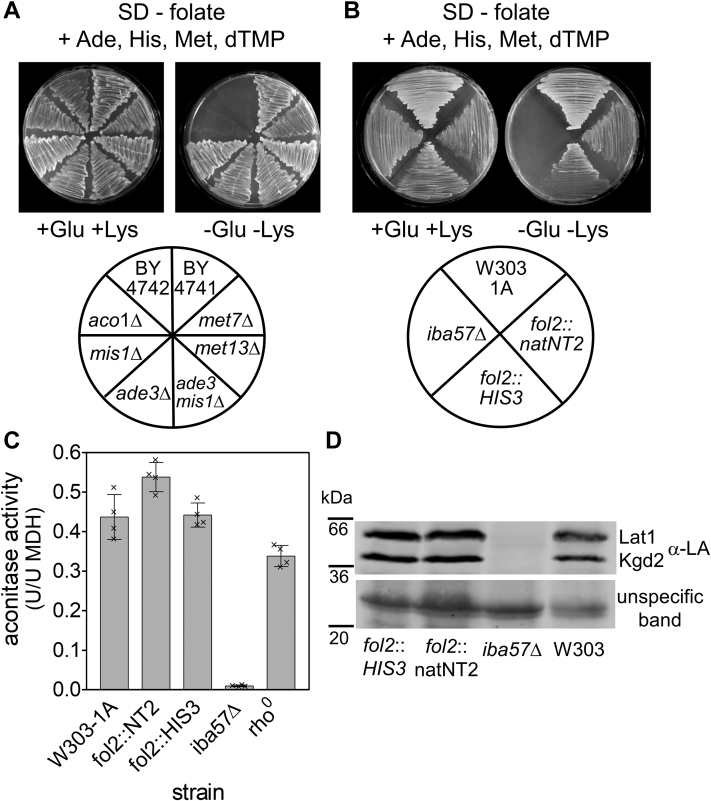
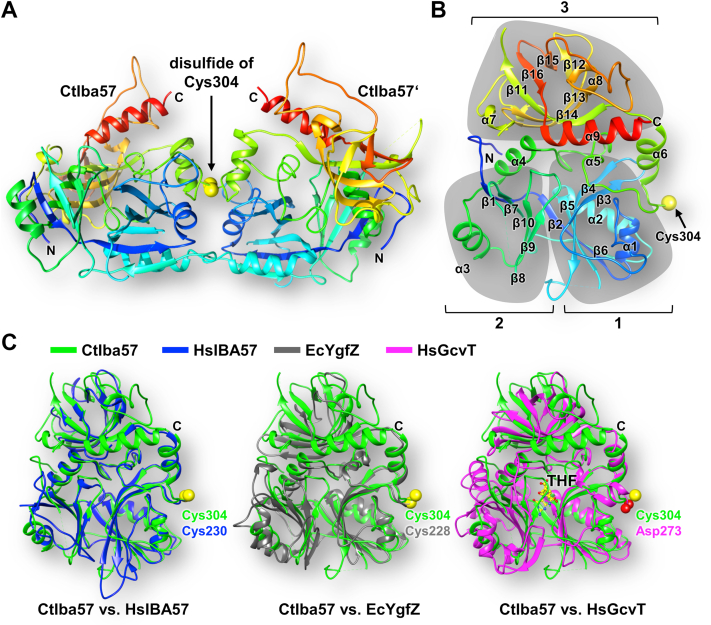
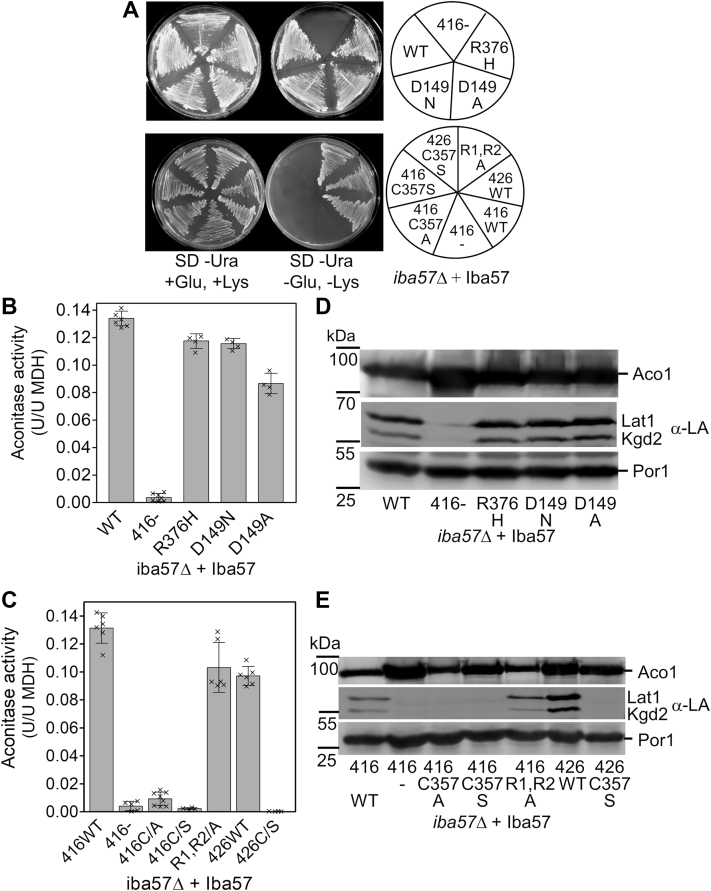
Mitochondria harbor the bacteria-inherited iron-sulfur cluster assembly (ISC) machinery to generate [2Fe-2S; iron-sulfur (Fe-S)] and [4Fe-4S] proteins. In yeast, assembly of [4Fe-4S] proteins specifically involves the ISC proteins Isa1, Isa2, Iba57, Bol3, and Nfu1. Functional defects in their human equivalents cause the multiple mitochondrial dysfunction syndromes, severe disorders with a broad clinical spectrum. The bacterial Iba57 ancestor YgfZ was described to require tetrahydrofolate (THF) for its function in the maturation of selected [4Fe-4S] proteins. Both YgfZ and Iba57 are structurally related to an enzyme family catalyzing THF-dependent one-carbon transfer reactions including GcvT of the glycine cleavage system. On this basis, a universally conserved folate requirement in ISC-dependent [4Fe-4S] protein biogenesis was proposed. To test this idea for mitochondrial Iba57, we performed genetic and biochemical studies in Saccharomyces cerevisiae, and we solved the crystal structure of Iba57 from the thermophilic fungus Chaetomium thermophilum. We provide three lines of evidence for the THF independence of the Iba57-catalyzed [4Fe-4S] protein assembly pathway. First, yeast mutants lacking folate show no defect in mitochondrial [4Fe-4S] protein maturation. Second, the 3D structure of Iba57 lacks many of the side-chain contacts to THF as defined in GcvT, and the THF-binding pocket is constricted. Third, mutations in conserved Iba57 residues that are essential for THF-dependent catalysis in GcvT do not impair Iba57 function in vivo, in contrast to an exchange of the invariant, surface-exposed cysteine residue. We conclude that mitochondrial Iba57, despite structural similarities to both YgfZ and THF-binding proteins, does not utilize folate for its function.
Keywords: crystallography; folate; iron–sulfur cluster assembly; iron–sulfur protein; mitochondria; mitochondrial disease.
Copyright © 2022 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures



References
-
- Zanello P. Structure and electrochemistry of proteins harboring iron-sulfur clusters of different nuclearities. Part IV. Canonical, non-canonical and hybrid iron-sulfur proteins. J. Struct. Biol. 2019;205:103–120. - PubMed
-
- Braymer J.J., Freibert S.A., Rakwalska-Bange M., Lill R. Mechanistic concepts of iron-sulfur protein biogenesis in biology. Biochim. Biophys. Acta Mol. Cell Res. 2021;1868 - PubMed
-
- Lill R., Freibert S.A. Mechanisms of mitochondrial iron-sulfur protein biogenesis. Annu. Rev. Biochem. 2020;89:471–499. - PubMed
-
- Ciofi-Baffoni S., Nasta V., Banci L. Protein networks in the maturation of human iron-sulfur proteins. Metallomics. 2018;10:49–72. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
