

Parasites and the neuroendocrine control of fish intestinal function: an ancient struggle between pathogens and host
- PMID: 36076315
- PMCID: PMC11010486
- DOI: 10.1017/S0031182022001160
Parasites and the neuroendocrine control of fish intestinal function: an ancient struggle between pathogens and host
Abstract
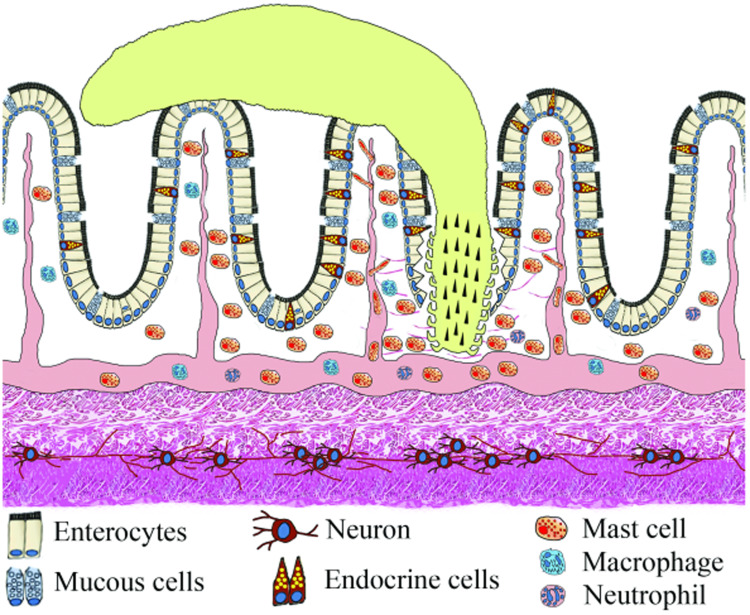
Most individual fish in wild and farmed populations can be infected with parasites. Fish intestines can harbour protozoans, myxozoans and helminths, which include several species of digeneans, cestodes, nematodes and acanthocephalans. Enteric parasites often induce inflammation of the intestine; the pathogen provokes changes in the host physiology, which will be genetically selected for if they benefit the parasite. The host response to intestinal parasites involves neural, endocrine and immune systems and interaction among these systems is coordinated by hormones, chemokines, cytokines and neurotransmitters including peptides. Intestinal fish parasites have effects on the components of the enteric nervous and endocrine systems; mechanical/chemical changes impair the activity of these systems, including gut motility and digestion. Investigations on the role of the neuroendocrine system in response to fish intestinal parasites are very few. This paper provides immunohistochemical and ultrastructural data on effects of parasites on the enteric nervous system and the enteric endocrine system in several fish–parasite systems. Emphasis is on the occurrence of 21 molecules including cholecystokinin-8, neuropeptide Y, enkephalins, galanin, vasoactive intestinal peptide and serotonin in infected tissues.
Keywords: Enteric endocrine system; enteric nervous system; gut physiology; intestinal parasites; teleost.
Conflict of interest statement
The authors declare having no conflict of interest.
Figures










Similar articles
-
Pike intestinal reaction to Acanthocephalus lucii (Acanthocephala): immunohistochemical and ultrastructural surveys.Parasit Vectors. 2018 Jul 16;11(1):424. doi: 10.1186/s13071-018-3002-6. Parasit Vectors. 2018. PMID: 30012189 Free PMC article.
-
Fish innate immunity against intestinal helminths.Fish Shellfish Immunol. 2016 Mar;50:274-87. doi: 10.1016/j.fsi.2016.02.002. Epub 2016 Feb 9. Fish Shellfish Immunol. 2016. PMID: 26868213 Review.
-
Enteric neuromodulators and mucus discharge in a fish infected with the intestinal helminth Pomphorhynchus laevis.Parasit Vectors. 2015 Jul 8;8:359. doi: 10.1186/s13071-015-0970-7. Parasit Vectors. 2015. PMID: 26152567 Free PMC article.
-
Marine fish parasites in the Cat Ba Archipelago, Vietnam: the results of 2010-2023 field surveys.Syst Parasitol. 2024 Feb 14;101(2):20. doi: 10.1007/s11230-023-10145-w. Syst Parasitol. 2024. PMID: 38351396
-
Survival of metazoan parasites in fish: Putting into context the protective immune responses of teleost fish.Adv Parasitol. 2021;112:77-132. doi: 10.1016/bs.apar.2021.03.001. Epub 2021 Apr 29. Adv Parasitol. 2021. PMID: 34024360 Review.
Cited by
-
Immunomodulatory and antioxidant responses of African catfish (Clarias gariepinus) to cestode infections: implications for intestinal health and aquaculture management.Vet Res Commun. 2025 Jul 11;49(5):254. doi: 10.1007/s11259-025-10819-4. Vet Res Commun. 2025. PMID: 40643779
-
Fish parasites (special issue).Parasitology. 2022 Dec;149(14):1811-1814. doi: 10.1017/S0031182022001433. Epub 2022 Oct 13. Parasitology. 2022. PMID: 36226653 Free PMC article.
-
Nitric Oxide Function and Nitric Oxide Synthase Evolution in Aquatic Chordates.Int J Mol Sci. 2023 Jul 6;24(13):11182. doi: 10.3390/ijms241311182. Int J Mol Sci. 2023. PMID: 37446358 Free PMC article. Review.
-
Hooking the scientific community on thorny-headed worms: interesting and exciting facts, knowledge gaps and perspectives for research directions on Acanthocephala.Parasite. 2023;30:23. doi: 10.1051/parasite/2023026. Epub 2023 Jun 22. Parasite. 2023. PMID: 37350678 Free PMC article.
-
Infection of endemic chub Squalius tenellus with the intestinal tapeworm Caryophyllaeus brachycollis (Cestoda): histopathology and ultrastructural surveys.Parasitology. 2024 Feb;151(2):157-167. doi: 10.1017/S0031182023001233. Epub 2023 Dec 1. Parasitology. 2024. PMID: 38193283 Free PMC article.
References
-
- Adamo SA (2002) Modulating the modulators: parasites, neuromodulators and host behavioural change. Brain Behavior and Evolution 60, 370–377. - PubMed
-
- Ahlman H and Nilsson O (2001) The gut as the largest endocrine organ in the body. Annals of Oncology 12, S63–S68. - PubMed
-
- Aldegunde M and Mancebo M (2006) Effects of neuropeptide Y on food intake and brain biogenic amines in the rainbow trout (Oncorhynchus mykiss). Peptides 27, 719–727. - PubMed
-
- Aldman G and Holmgren S (1995) Intraduodenal fat and amino acids activate gallbladder motility in the rainbow trout, Oncorhynchus mykiss. General and Comparative Endocrinology 100, 27–32. - PubMed
-
- Anderson RC (2000) Nematode Parasites of Vertebrates: Their Development and Transmission. Wallingford, UK: CABI Publishing.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
