

Chlamydomonas reinhardtii Alternates Peroxisomal Contents in Response to Trophic Conditions
- PMID: 36078132
- PMCID: PMC9454557
- DOI: 10.3390/cells11172724
Chlamydomonas reinhardtii Alternates Peroxisomal Contents in Response to Trophic Conditions
Abstract
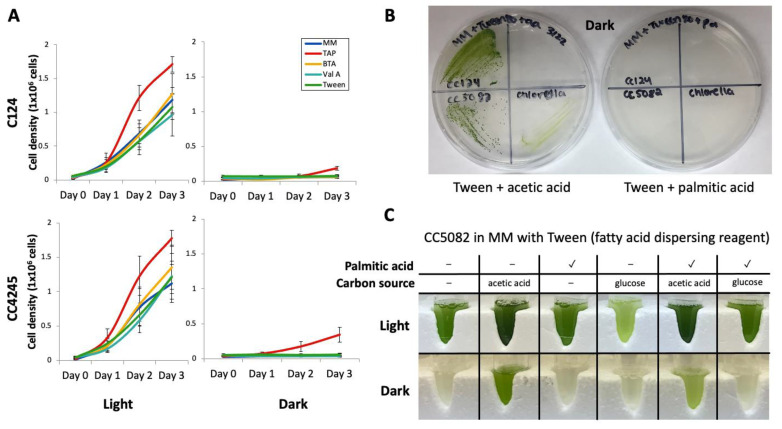
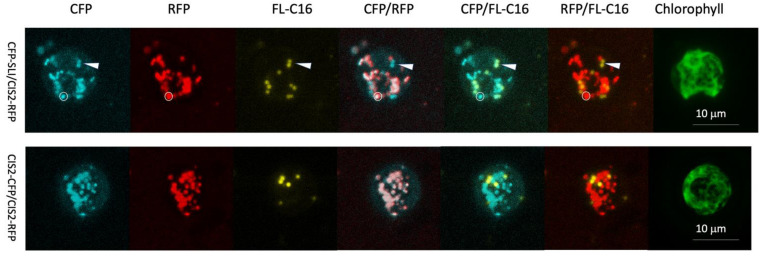
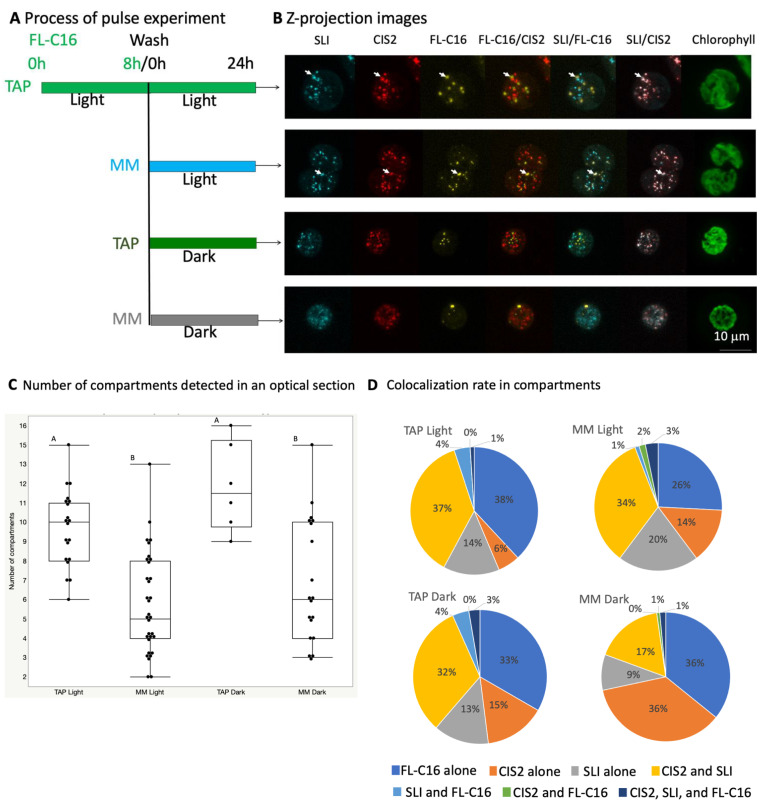
Chlamydomonas reinhardtii is a model green microalga capable of heterotrophic growth on acetic acid but not fatty acids, despite containing a full complement of genes for β-oxidation. Recent reports indicate that the alga preferentially sequesters, rather than breaks down, lipid acyl chains as a means to rebuild its membranes rapidly. Here, we assemble a list of potential Chlamydomonas peroxins (PEXs) required for peroxisomal biogenesis to suggest that C. reinhardtii has a complete set of peroxisome biogenesis factors. To determine involvements of the peroxisomes in the metabolism of exogenously added fatty acids, we examined transgenic C. reinhardtii expressing fluorescent proteins fused to N- or C-terminal peptide of peroxisomal proteins, concomitantly with fluorescently labeled palmitic acid under different trophic conditions. We used confocal microscopy to track the populations of the peroxisomes in illuminated and dark conditions, with and without acetic acid as a carbon source. In the cells, four major populations of compartments were identified, containing: (1) a glyoxylate cycle enzyme marker and a protein containing peroxisomal targeting signal 1 (PTS1) tripeptide but lacking the fatty acid marker, (2) the fatty acid marker alone, (3) the glyoxylate cycle enzyme marker alone, and (4) the PTS1 marker alone. Less than 5% of the compartments contained both fatty acid and peroxisomal markers. Statistical analysis on optically sectioned images found that C. reinhardtii simultaneously carries diverse populations of the peroxisomes in the cell and modulates peroxisomal contents based on light conditions. On the other hand, the ratio of the compartment containing both fatty acid and peroxisomal markers did not change significantly regardless of the culture conditions. The result indicates that β-oxidation may be only a minor occurrence in the peroxisomal population in C. reinhardtii, which supports the idea that lipid biosynthesis and not β-oxidation is the primary metabolic preference of fatty acids in the alga.
Keywords: Chlamydomonas; PEX; PTS1; fatty acids; fluorescence microscopy; glyoxylate cycle enzymes; microalgae; peroxisomes; β-oxidation.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
