

Na,K-ATPase Acts as a Beta-Amyloid Receptor Triggering Src Kinase Activation
- PMID: 36078160
- PMCID: PMC9455167
- DOI: 10.3390/cells11172753
Na,K-ATPase Acts as a Beta-Amyloid Receptor Triggering Src Kinase Activation
Abstract
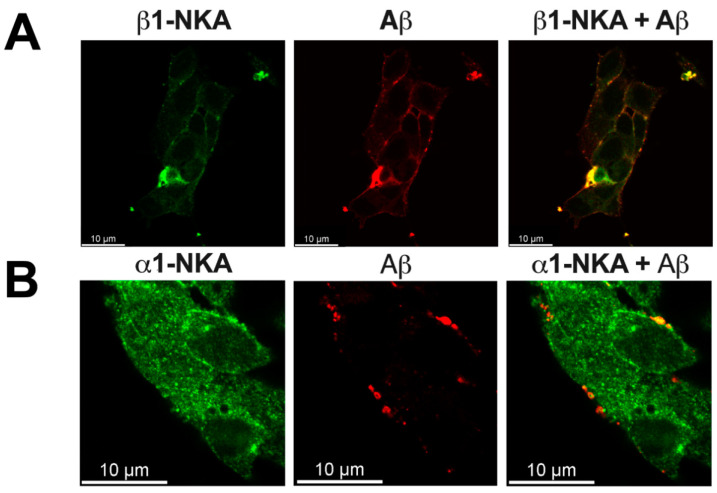
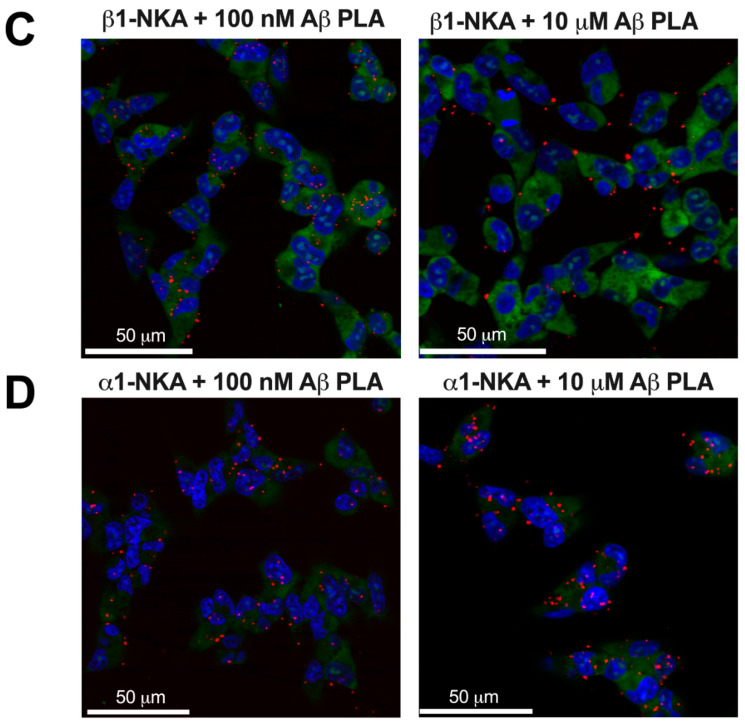
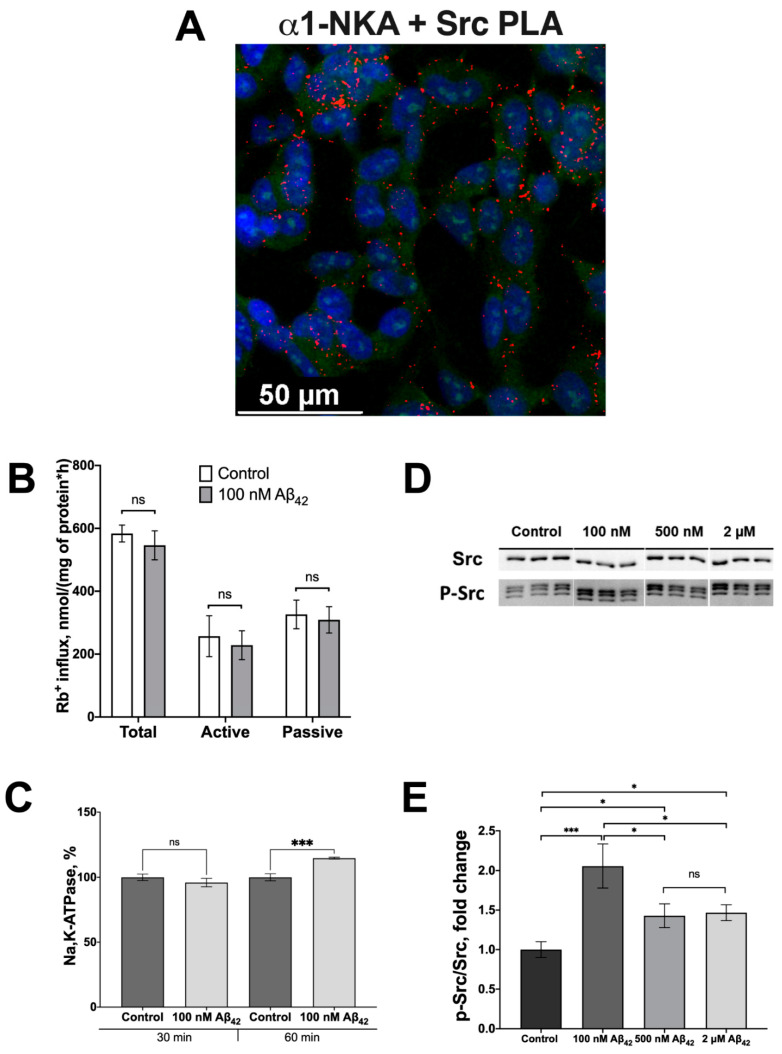
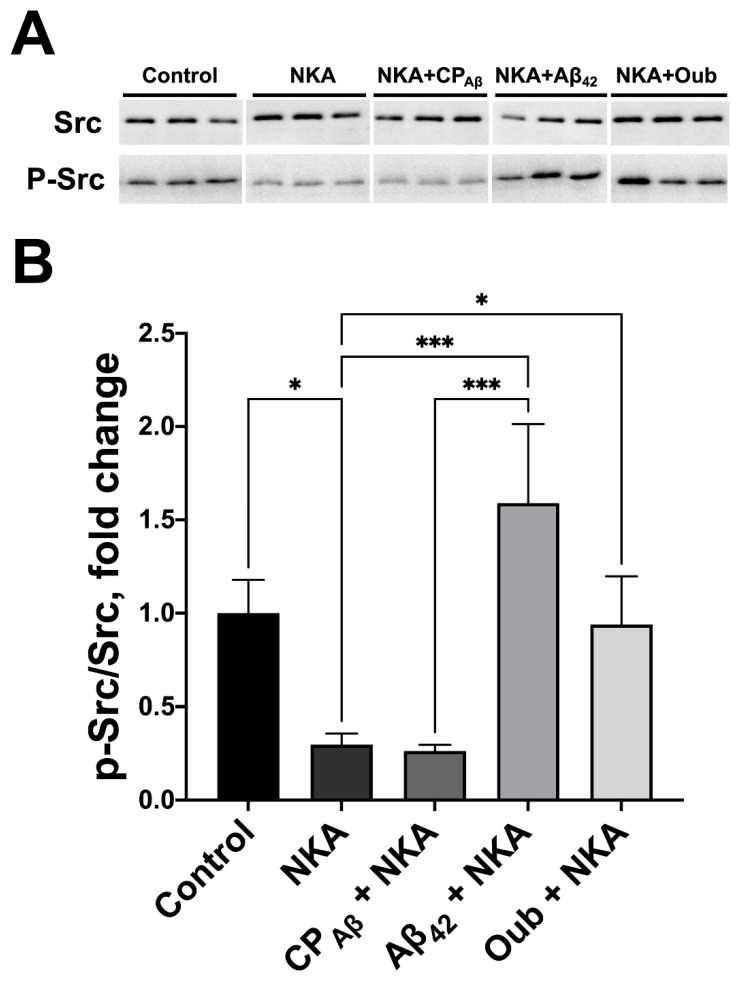
Beta-amyloid (Aβ) has a dual role, both as an important factor in the pathology of Alzheimer's disease and as a regulator in brain physiology. The inhibitory effect of Aβ42 oligomers on Na,K-ATPase contributes to neuronal dysfunction in Alzheimer's disease. Still, the physiological role of the monomeric form of Aβ42 interaction with Na,K-ATPase remains unclear. We report that Na,K-ATPase serves as a receptor for Aβ42 monomer, triggering Src kinase activation. The co-localization of Aβ42 with α1- and β1-subunits of Na,K-ATPase, and Na,K-ATPase with Src kinase in SH-SY5Y neuroblastoma cells, was observed. Treatment of cells with 100 nM Aβ42 causes Src kinase activation, but does not alter Na,K-ATPase transport activity. The interaction of Aβ42 with α1β1 Na,K-ATPase isozyme leads to activation of Src kinase associated with the enzyme. Notably, prevention of Na,K-ATPase:Src kinase interaction by a specific inhibitor pNaKtide disrupts the Aβ-induced Src kinase activation. Stimulatory effect of Aβ42 on Src kinase was lost under hypoxic conditions, which was similar to the effect of specific Na,K-ATPase ligands, the cardiotonic steroids. Our findings identify Na,K-ATPase as a Aβ42 receptor, thus opening a prospect on exploring the physiological and pathological Src kinase activation caused by Aβ42 in the nervous system.
Keywords: Na,K-ATPase; Src kinase; beta-amyloid; hypoxia; receptor function.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
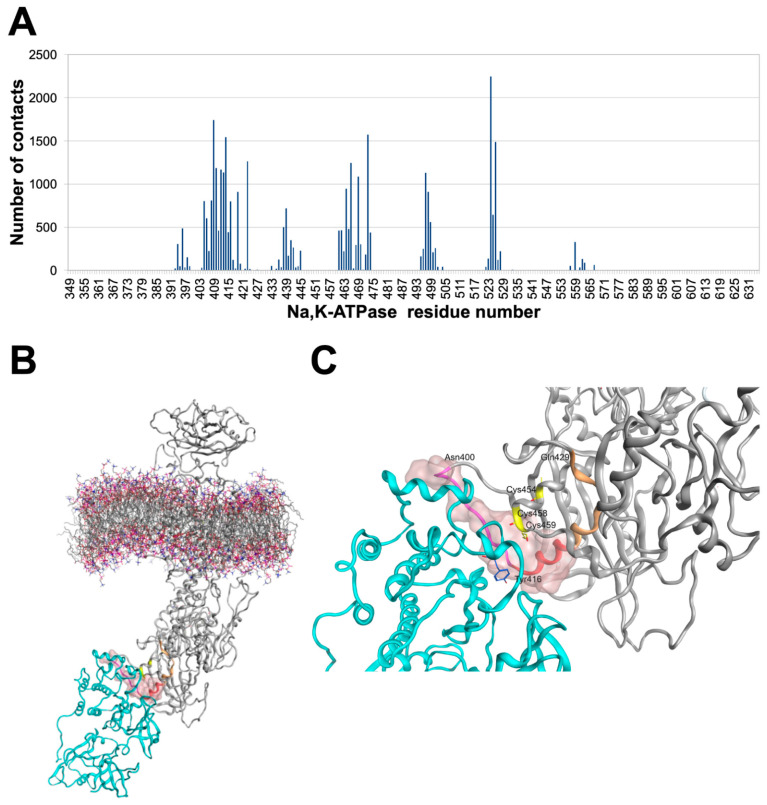
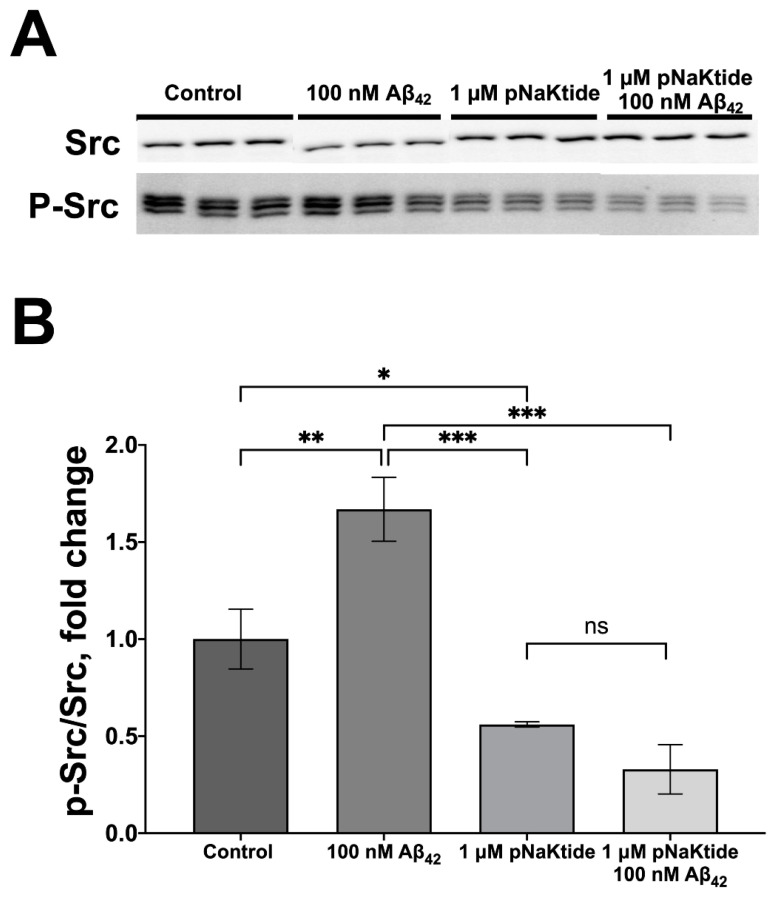
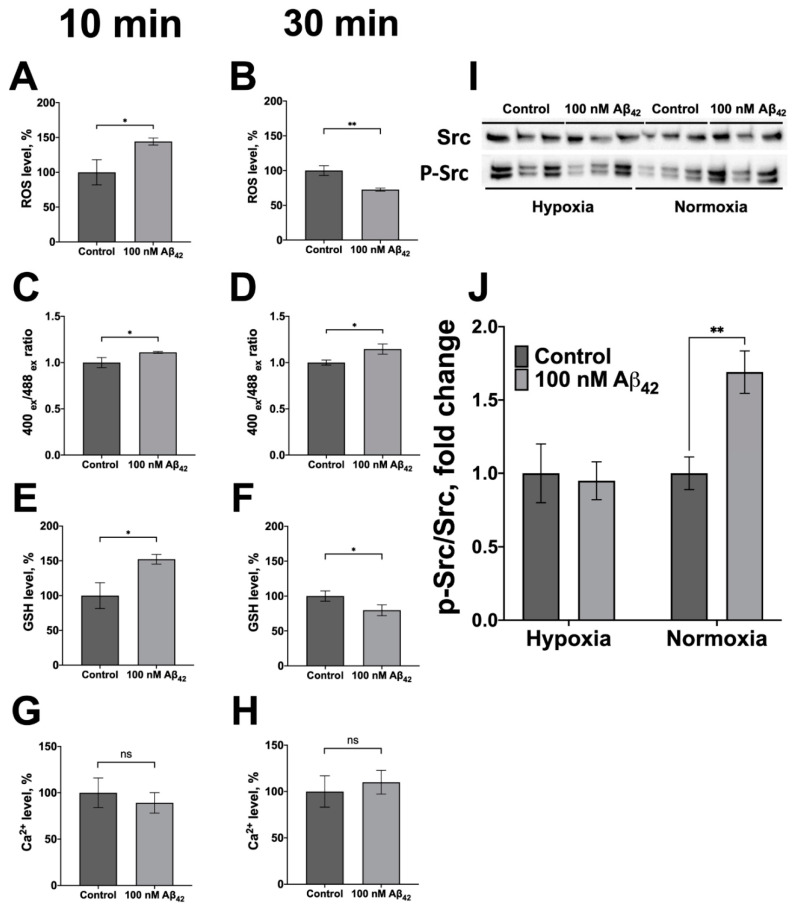
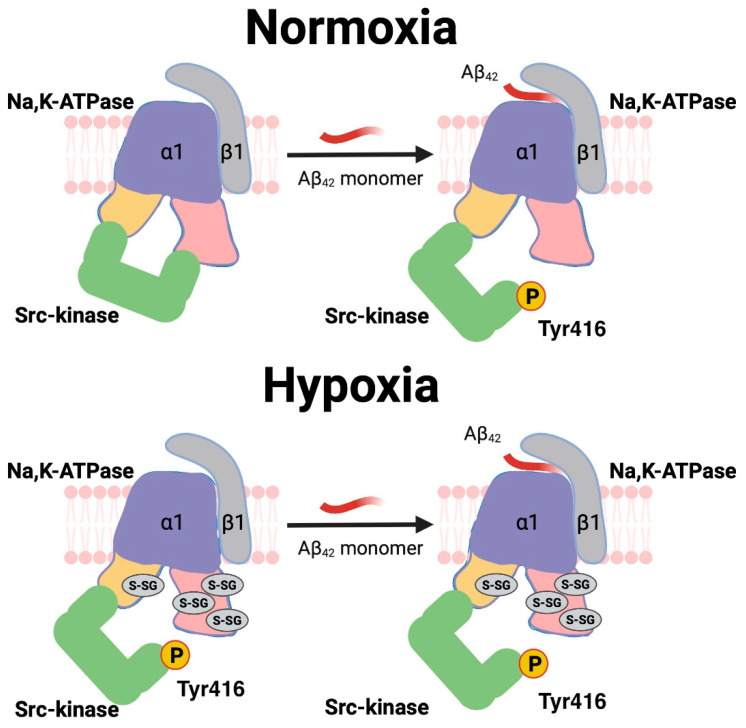
References
-
- Meyer-Luehmann M., Coomaraswamy J., Bolmont T., Kaeser S., Schaefer C., Kilger E., Neuenschwander A., Abramowski D., Frey P., Jaton A.L., et al. Exogenous induction of cerebral beta-amyloidogenesis is governed by agent and host. Science. 2006;313:1781–1784. doi: 10.1126/science.1131864. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
