

Manipulation of mRNA translation elongation influences the fragmentation of a biotherapeutic Fc-fusion protein produced in CHO cells
- PMID: 36082414
- PMCID: PMC9826484
- DOI: 10.1002/bit.28230
Manipulation of mRNA translation elongation influences the fragmentation of a biotherapeutic Fc-fusion protein produced in CHO cells
Abstract
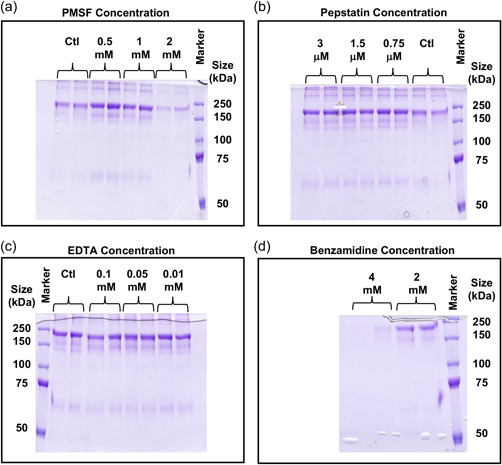
Mammalian cells, particularly Chinese hamster ovary cells, are the dominant system for the production of protein-based biotherapeutics, however, product degradation, particularly of Fc-fusion proteins, is sometimes observed that impacts the quality of the protein generated. Here, we identify the site of fragmentation of a model immunoglobulin G1 Fc-fusion protein, show that the observed clipping and aggregation are decreased by reduced temperature culturing, that the fragmentation/clipping is intracellular, and that reduced clipping at a lower temperature (<37°C) relates to mesenger RNA (mRNA) translation elongation. We subsequently show that reduced fragmentation can be achieved at 37°C by addition of chemical reagents that slow translation elongation. We then modified mRNA translation elongation speeds by designing different transcript sequences for the Fc-fusion protein based on alternative codon usage and improved the product yield at 37°C, and the ratio of intact to a fragmented product. Our data suggest that rapid elongation results in misfolding that decreases product fidelity, generating a region susceptible to degradation/proteolysis, whilst the slowing of mRNA translation improves the folding, reducing susceptibility to fragmentation. Manipulation of mRNA translation and/or the target Fc-fusion transcript is, therefore, an approach that can be applied to potentially reduce fragmentation of clipping-prone Fc-fusion proteins.
Keywords: Chinese hamster ovary (CHO) cells; Fc-fusion protein; clipping; fragmentation; mRNA translation elongation.
© 2022 The Authors. Biotechnology and Bioengineering published by Wiley Periodicals LLC.
Conflict of interest statement
Atul Mohindra and Colin M. Jaques are employed by Lonza Biologics. Lonza Biologics owns and licenses the GS Gene Expression System®, CHOK1SV®, and CHOK1SV GS‐KO® cell lines. The remaining authors declare no conflict of interest.
Figures






Similar articles
-
The Effect of Culture Temperature on the Aggregation of Recombinant TNFR-Fc is Regulated by the PERK-eIF2a Pathway in CHO Cells.Protein Pept Lett. 2018;25(6):570-579. doi: 10.2174/0929866525666180530121317. Protein Pept Lett. 2018. PMID: 29848262
-
Understanding the intracellular effects of yeast extract on the enhancement of Fc-fusion protein production in Chinese hamster ovary cell culture.Appl Microbiol Biotechnol. 2015 Oct;99(20):8429-40. doi: 10.1007/s00253-015-6789-5. Epub 2015 Jul 11. Appl Microbiol Biotechnol. 2015. PMID: 26162671
-
Studies to Prevent Degradation of Recombinant Fc-Fusion Protein Expressed in Mammalian Cell Line and Protein Characterization.Int J Mol Sci. 2016 Jun 9;17(6):913. doi: 10.3390/ijms17060913. Int J Mol Sci. 2016. PMID: 27294920 Free PMC article.
-
Full-length recombinant antibodies from Escherichia coli: production, characterization, effector function (Fc) engineering, and clinical evaluation.MAbs. 2022 Jan-Dec;14(1):2111748. doi: 10.1080/19420862.2022.2111748. MAbs. 2022. PMID: 36018829 Free PMC article. Review.
-
Mammalian cell-produced therapeutic proteins: heterogeneity derived from protein degradation.Curr Opin Biotechnol. 2014 Dec;30:198-204. doi: 10.1016/j.copbio.2014.07.007. Epub 2014 Aug 12. Curr Opin Biotechnol. 2014. PMID: 25118137 Review.
Cited by
-
Recombinant therapeutic proteins degradation and overcoming strategies in CHO cells.Appl Microbiol Biotechnol. 2024 Jan 29;108(1):182. doi: 10.1007/s00253-024-13008-6. Appl Microbiol Biotechnol. 2024. PMID: 38285115 Free PMC article. Review.
References
-
- Bryan, L. , Henry, H. , Barron, N. , Gallagher, C. , Kelly, R. M. , Frye, C. C. , Osborne, M. D. , Clynes, M. , Meleady, P. (2021). Differential expression of miRNAs and functional role of miR‐200a in high and low productivity CHO cells expressing an Fc fusion protein, Biotechnology Letters, 43, 1551–1563. 10.1007/s10529-021-03153-7 - DOI - PMC - PubMed
-
- Budge, J. D. , Knight, T. J. , Povey, J. , Roobol, J. , Brown, I. R. , Singh, G. , Dean, A. , Turner, S. , Jaques, C. M. , Young, R. J. , Racher, A. J. , Smales, C. M. (2020). Engineering of Chinese hamster ovary cell lipid metabolism results in an expanded ER and enhanced recombinant biotherapeutic protein production, Metabolic Engineering, 57, 203–216. 10.1016/j.ymben.2019.11.007 - DOI - PMC - PubMed
-
- Budge, J. D. , Roobol, J. , Singh, G. , Mozzanino, T. , Knight, T. J. , Povy, J. , Dean, A. , Turner, S. J. , Jaques, C. M. , Young, R. J. , Racher, A. J. , Smales, C. M. (2021). A proline metabolism selection system and its application to the engineering of lipid biosynthesis in Chinese hamster ovary cells, Metabolic Engineering Communications, 13, e00179. 10.1016/j.mec.2021.e00179 - DOI - PMC - PubMed
-
- Chakrabarti, S. , Barrow C. J., Kanwar R. K., Ramana V., Kanwar J. R. (2016). ‘Studies to prevent degradation of recombinant Fc‐fusion protein expressed in mammalian cell line and protein characterization’, International Journal of Molecular Sciences, 17(6), 913. 10.3390/ijms17060913 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
