

Integrated changes in thermal stability and proteome abundance during altered nutrient states in Escherichia coli and human cells
- PMID: 36082775
- PMCID: PMC10782824
- DOI: 10.1002/pmic.202100254
Integrated changes in thermal stability and proteome abundance during altered nutrient states in Escherichia coli and human cells
Abstract
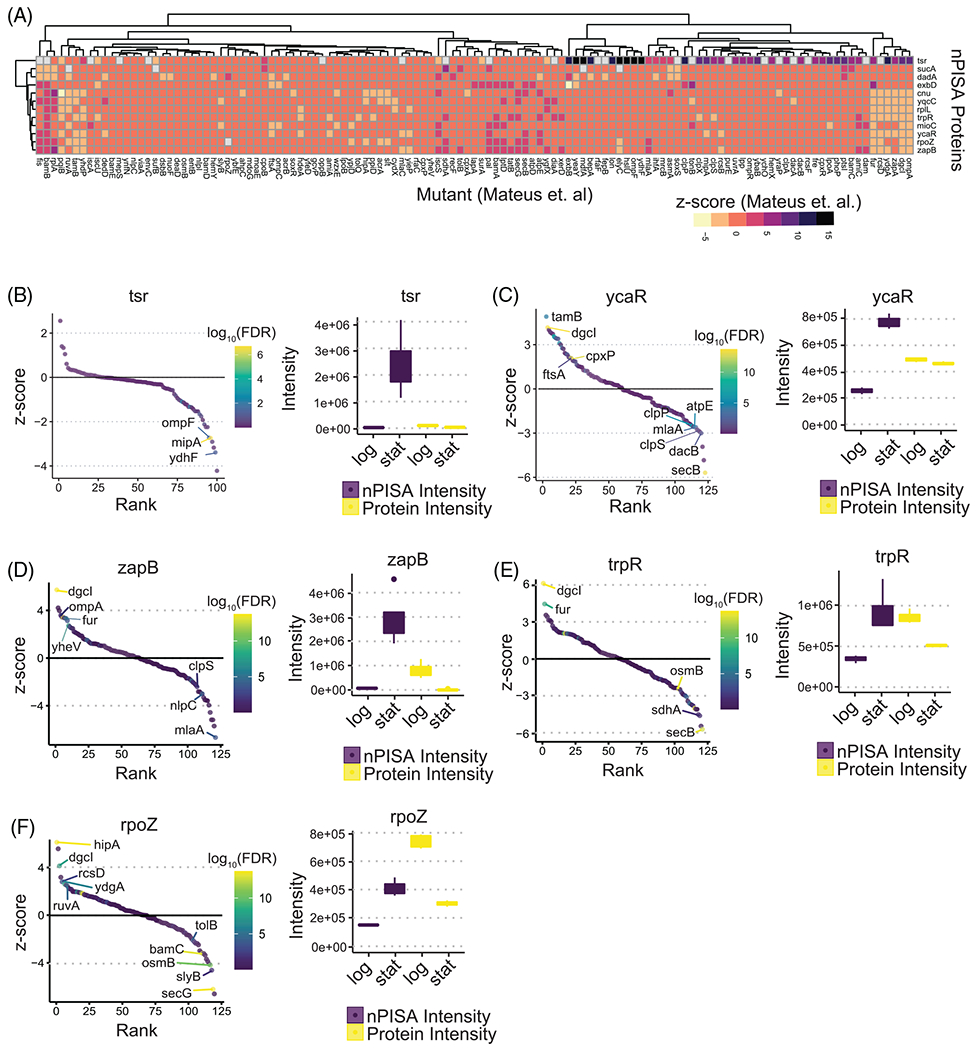
Altered thermal solubility measurement techniques are emerging as powerful tools to assess ligand binding, post-translational modification, protein-protein interactions, and many other cellular processes that affect protein state under various cellular conditions. Thermal solubility or stability profiling techniques are enabled on a global proteomic scale by employing isobaric tagging reagents that facilitate multiplexing capacity required to measure changes in the proteome across thermal gradients. Key among these is thermal proteomic profiling (TPP), which requires 8-10 isobaric tags per gradient and generation of multiple proteomic datasets to measure different replicates and conditions. Furthermore, using TPP to measure protein thermal stability state across different conditions may also require measurements of differential protein abundance. Here, we use the proteome integral stability alteration (PISA) assay, a higher throughput version of TPP, to measure global changes in protein thermal stability normalized to their protein abundance. We explore the use of this approach to determine changes in protein state between logarithmic and stationary phase Escherichia coli as well as glucose-starved human Hek293T cells. We observed protein intensity-corrected PISA changes in 290 and 350 proteins due to stationary phase transition in E. coli and glucose starvation, respectively. These data reveal several examples of proteins that were not previously associated with nutrient states by abundance alone. These include E. coli proteins such as putative acyl-CoA dehydrogenase (aidB) and chaperedoxin (cnoX) as well as human RAB vesicle trafficking proteins and many others which may indicate their involvement in metabolic diseases such as cancer.
Keywords: E. coli stationary phase; glucose starvation; isobaric peptide tagging; ligand-protein interactions; proteome integral stability alteration assay.
© 2022 Wiley-VCH GmbH.
Conflict of interest statement
CONFLICT OF INTEREST
The authors declare no conflict of interest.
Figures





References
-
- Molina DM, Jafari R, Ignatushchenko M, Seki T, Larsson EA, Dan C, Sreekumar L, Cao Y, & Nordlund P (2013). Monitoring drug target engagement in cells and tissues using the cellular thermal shift assay. Science, 341, 84–87. - PubMed
-
- Franken H, Mathieson T, Childs D, Sweetman GMA, Werner T, Tögel I, Doce C, Gade S, Bantscheff M, Drewes G, Reinhard FBM, Huber W, & Savitski MM (2015). Thermal proteome profiling for unbiased identification of direct and indirect drug targets using multiplexed quantitative mass spectrometry. Nature Protocols, 10, 1567–1593. - PubMed
-
- Savitski MM, Reinhard FBM, Franken H, Werner T, Savitski MF, Eberhard D, Molina DM, Jafari R, Dovega RB, Klaeger S, Kuster B, Nordlund P, Bantscheff M, & Drewes G (2014). Tracking cancer drugs in living cells by thermal profiling of the proteome. Science, 346, 1255784. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
