CtIP-dependent nascent RNA expression flanking DNA breaks guides the choice of DNA repair pathway
- PMID: 36085345
- PMCID: PMC9463442
- DOI: 10.1038/s41467-022-33027-z
CtIP-dependent nascent RNA expression flanking DNA breaks guides the choice of DNA repair pathway
Abstract
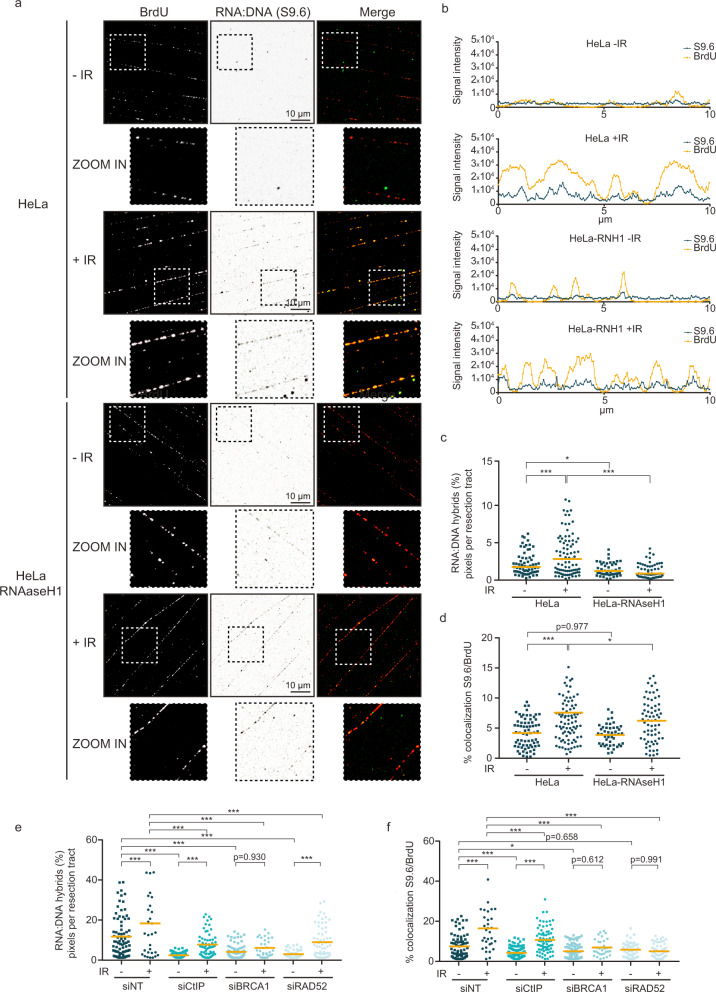
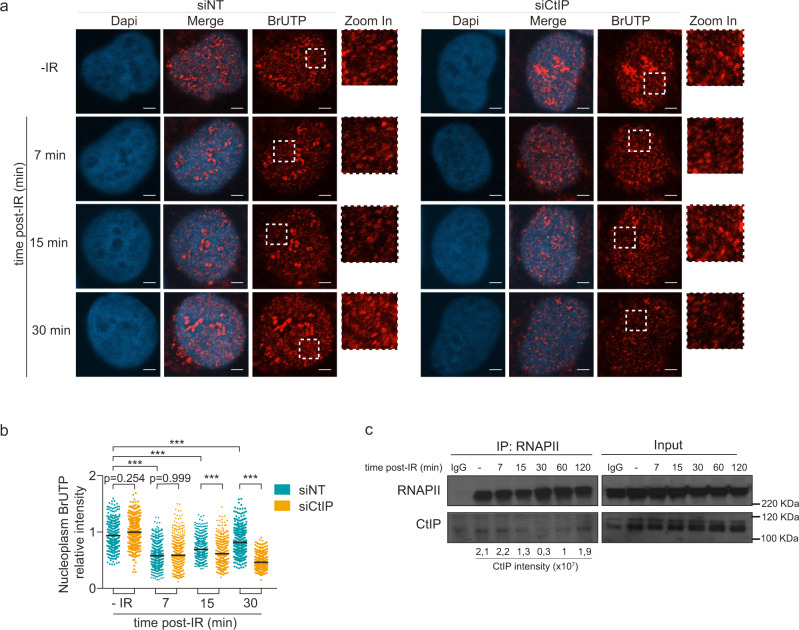
The RNA world is changing our views about sensing and resolution of DNA damage. Here, we develop single-molecule DNA/RNA analysis approaches to visualize how nascent RNA facilitates the repair of DNA double-strand breaks (DSBs). RNA polymerase II (RNAPII) is crucial for DSB resolution in human cells. DSB-flanking, RNAPII-generated nascent RNA forms RNA:DNA hybrids, guiding the upstream DNA repair steps towards favouring the error-free Homologous Recombination (HR) pathway over Non-Homologous End Joining. Specific RNAPII inhibitor, THZ1, impairs recruitment of essential HR proteins to DSBs, implicating nascent RNA in DNA end resection, initiation and execution of HR repair. We further propose that resection factor CtIP interacts with and helps re-activate RNAPII when paused by the RNA:DNA hybrids, collectively promoting faithful repair of chromosome breaks to maintain genomic integrity.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures