

Coronarin A modulated hepatic glycogen synthesis and gluconeogenesis via inhibiting mTORC1/S6K1 signaling and ameliorated glucose homeostasis of diabetic mice
- PMID: 36085523
- PMCID: PMC9958036
- DOI: 10.1038/s41401-022-00985-5
Coronarin A modulated hepatic glycogen synthesis and gluconeogenesis via inhibiting mTORC1/S6K1 signaling and ameliorated glucose homeostasis of diabetic mice
Abstract
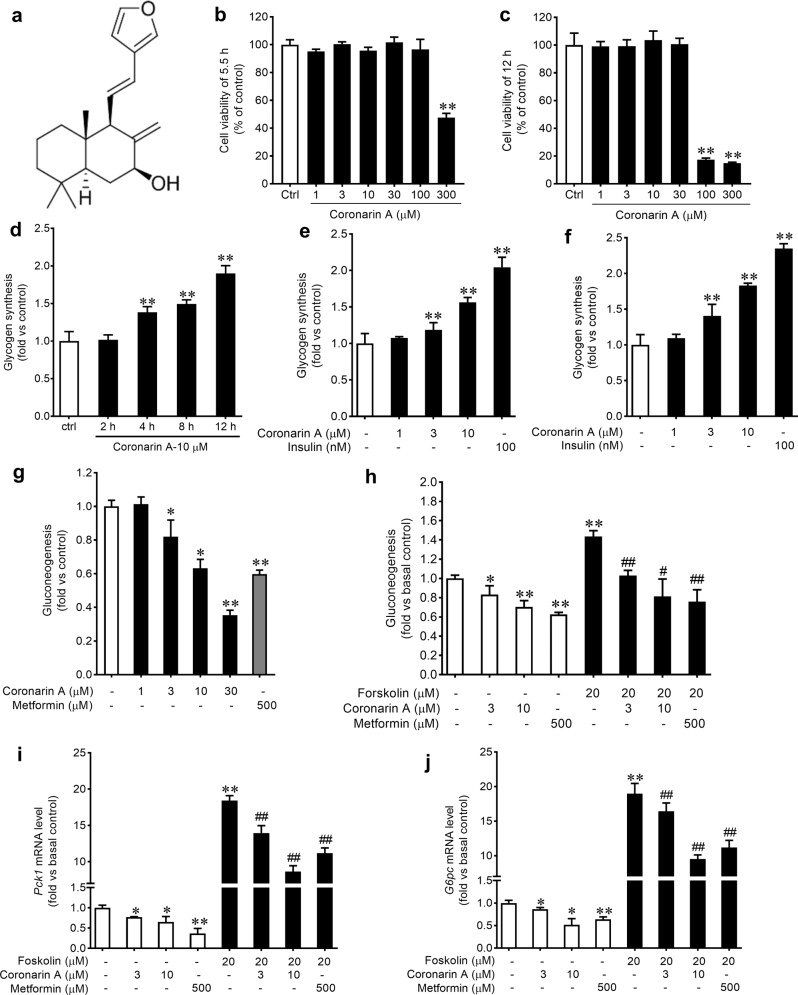
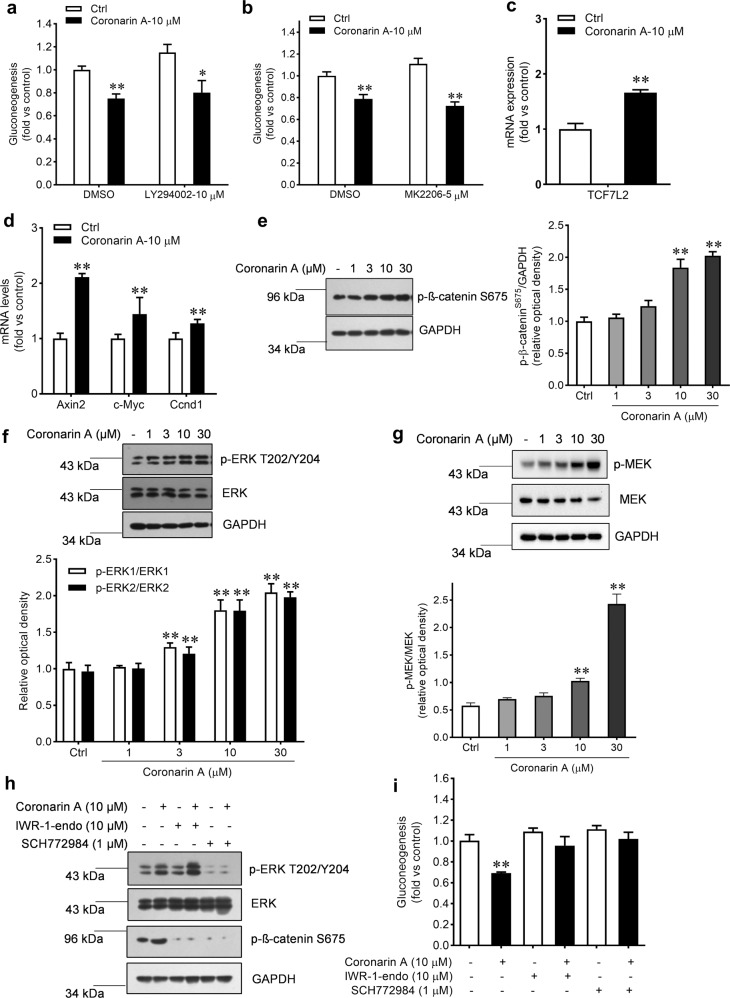
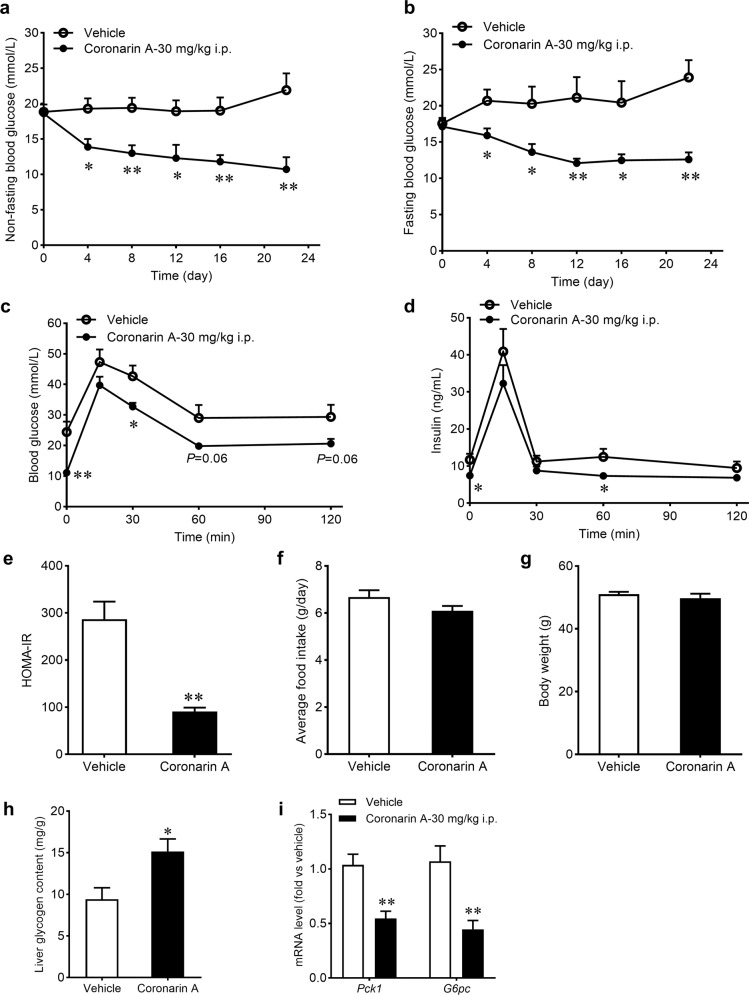
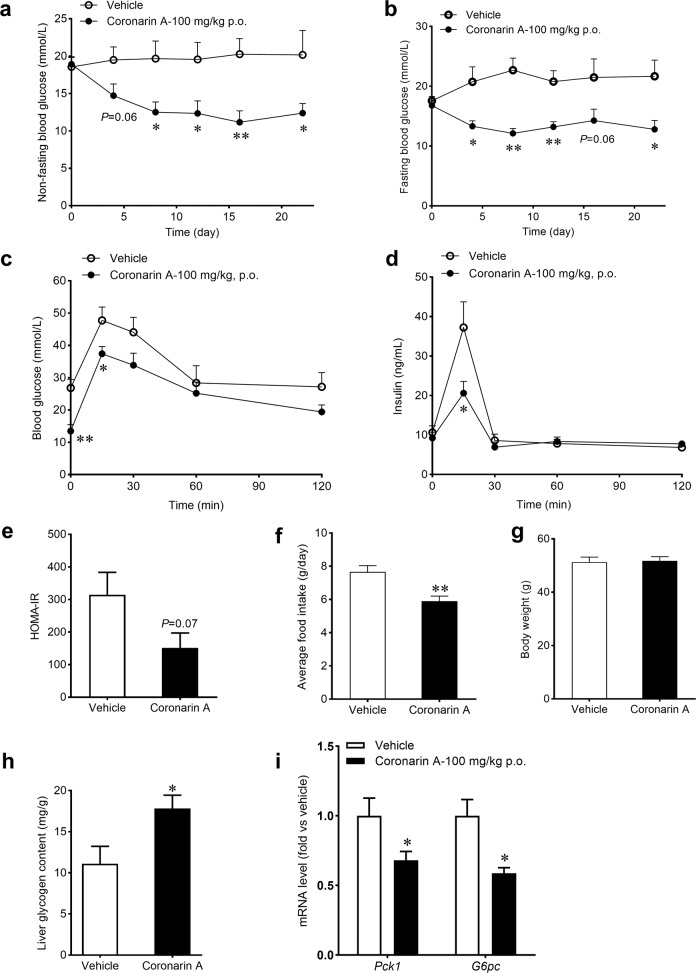
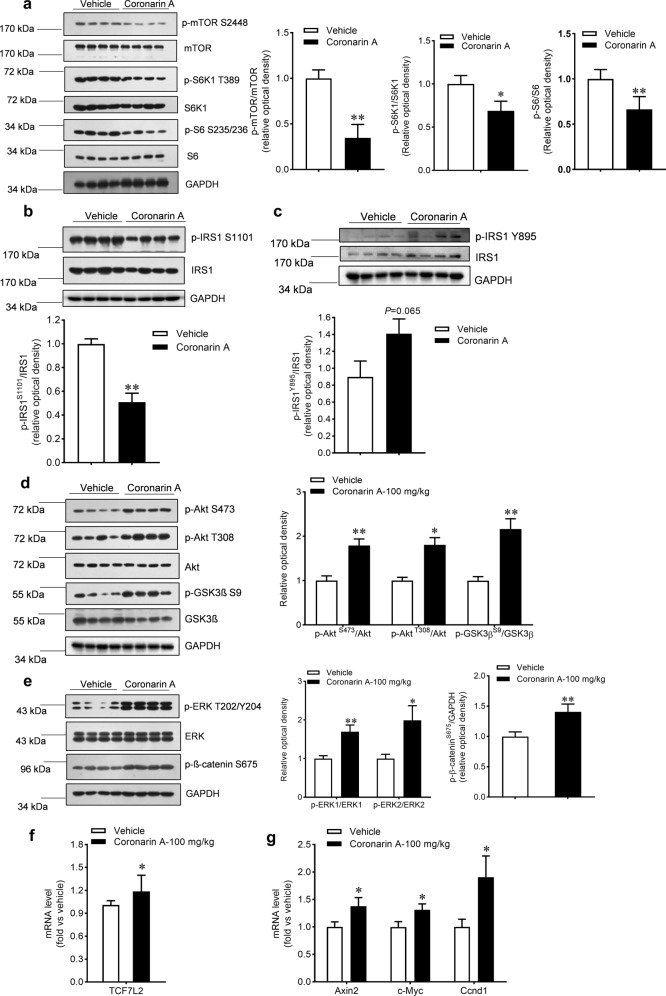
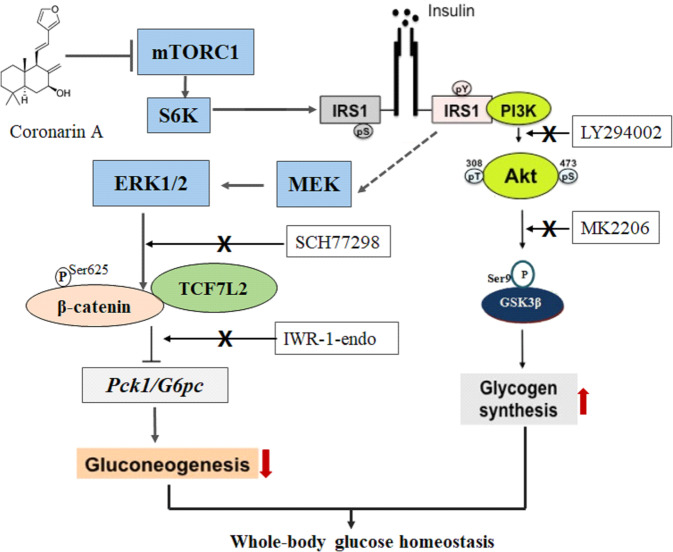
Promotion of hepatic glycogen synthesis and inhibition of hepatic glucose production are effective strategies for controlling hyperglycemia in type 2 diabetes mellitus (T2DM), but agents with both properties were limited. Herein we report coronarin A, a natural compound isolated from rhizomes of Hedychium gardnerianum, which simultaneously stimulates glycogen synthesis and suppresses gluconeogenesis in rat primary hepatocytes. We showed that coronarin A (3, 10 μM) dose-dependently stimulated glycogen synthesis accompanied by increased Akt and GSK3β phosphorylation in rat primary hepatocytes. Pretreatment with Akt inhibitor MK-2206 (2 μM) or PI3K inhibitor LY294002 (10 μM) blocked coronarin A-induced glycogen synthesis. Meanwhile, coronarin A (10 μM) significantly suppressed gluconeogenesis accompanied by increased phosphorylation of MEK, ERK1/2, β-catenin and increased the gene expression of TCF7L2 in rat primary hepatocytes. Pretreatment with β-catenin inhibitor IWR-1-endo (10 μM) or ERK inhibitor SCH772984 (1 μM) abolished the coronarin A-suppressed gluconeogenesis. More importantly, we revealed that coronarin A activated PI3K/Akt/GSK3β and ERK/Wnt/β-catenin signaling via regulation of a key upstream molecule IRS1. Coronarin A (10, 30 μM) decreased the phosphorylation of mTOR and S6K1, the downstream target of mTORC1, which further inhibited the serine phosphorylation of IRS1, and subsequently increased the tyrosine phosphorylation of IRS1. In type 2 diabetic ob/ob mice, chronic administration of coronarin A significantly reduced the non-fasting and fasting blood glucose levels and improved glucose tolerance, accompanied by the inhibited hepatic mTOR/S6K1 signaling and activated IRS1 along with enhanced PI3K/Akt/GSK3β and ERK/Wnt/β-catenin pathways. These results demonstrate the anti-hyperglycemic effect of coronarin A with a novel mechanism by inhibiting mTORC1/S6K1 to increase IRS1 activity, and highlighted coronarin A as a valuable lead compound for the treatment of T2DM.
Keywords: IRS1; coronarin A; gluconeogenesis; glycogen synthesis; mTORC1/S6K1 pathway; type 2 diabetes mellitus.
© 2022. The Author(s), under exclusive licence to Shanghai Institute of Materia Medica, Chinese Academy of Sciences and Chinese Pharmacological Society.
Conflict of interest statement
The authors declare no competing interests.
Figures


References
-
- Hwang JH, Perseghin G, Rothman DL, Cline GW, Magnusson I, Petersen KF, et al. Impaired net hepatic glycogen synthesis in insulin-dependent diabetic subjects during mixed meal ingestion. A 13C nuclear magnetic resonance spectroscopy study. J Clin Invest. 1995;95:783–7. doi: 10.1172/JCI117727. - DOI - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
