

Atractylenolide I ameliorates cancer cachexia through inhibiting biogenesis of IL-6 and tumour-derived extracellular vesicles
- PMID: 36085573
- PMCID: PMC9745491
- DOI: 10.1002/jcsm.13079
Atractylenolide I ameliorates cancer cachexia through inhibiting biogenesis of IL-6 and tumour-derived extracellular vesicles
Abstract
Background: Atractylenolide I (AI) is a natural sesquiterpene lactone isolated from Atractylodes macrocephala Koidz, known as Baizhu in traditional Chinese medicine. AI has been found to ameliorate cancer cachexia in clinic cancer patients and in tumour-bearing mice. Here, we checked the influence of AI on biogenesis of IL-6 and extracellular vesicles (EVs) in cancer cachexia mice and then focused on studying mechanisms of AI in inhibiting the production of tumour-derived EVs, which contribute to the ameliorating effects of AI on cancer cachexia.
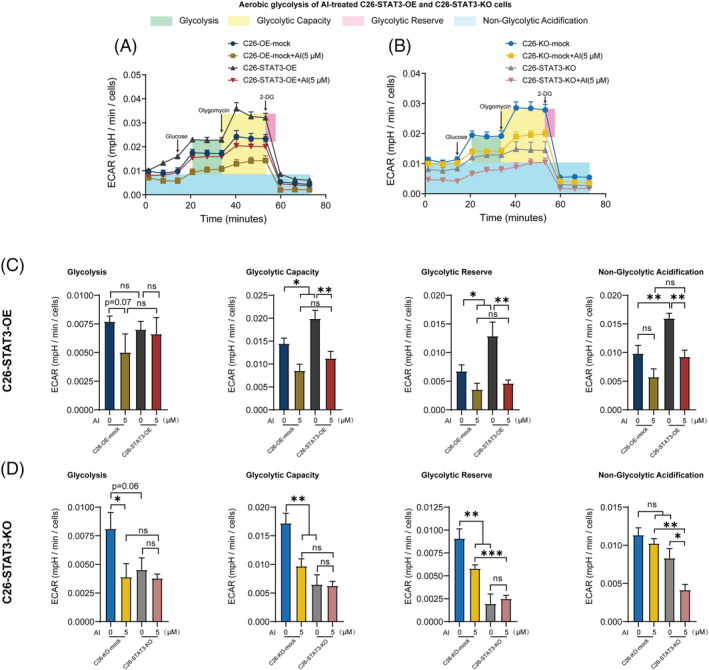
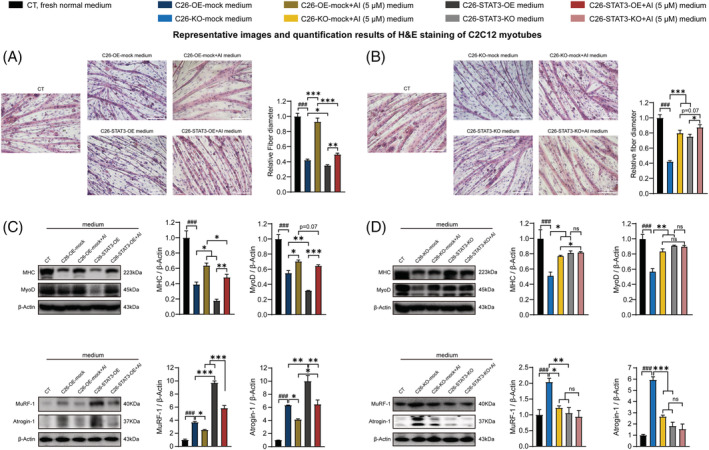
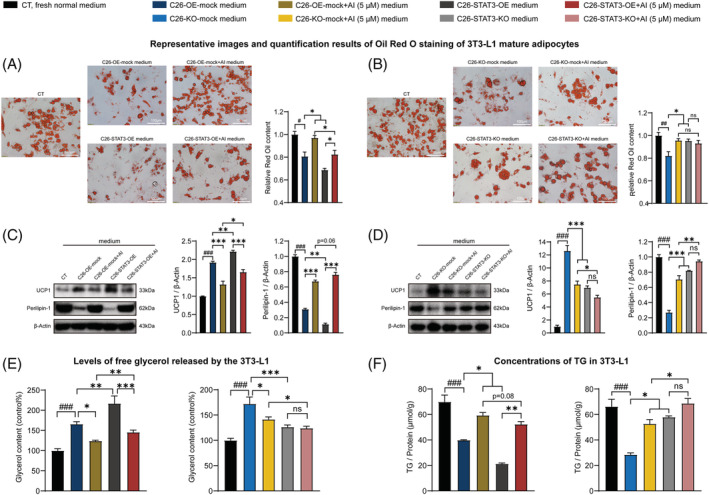
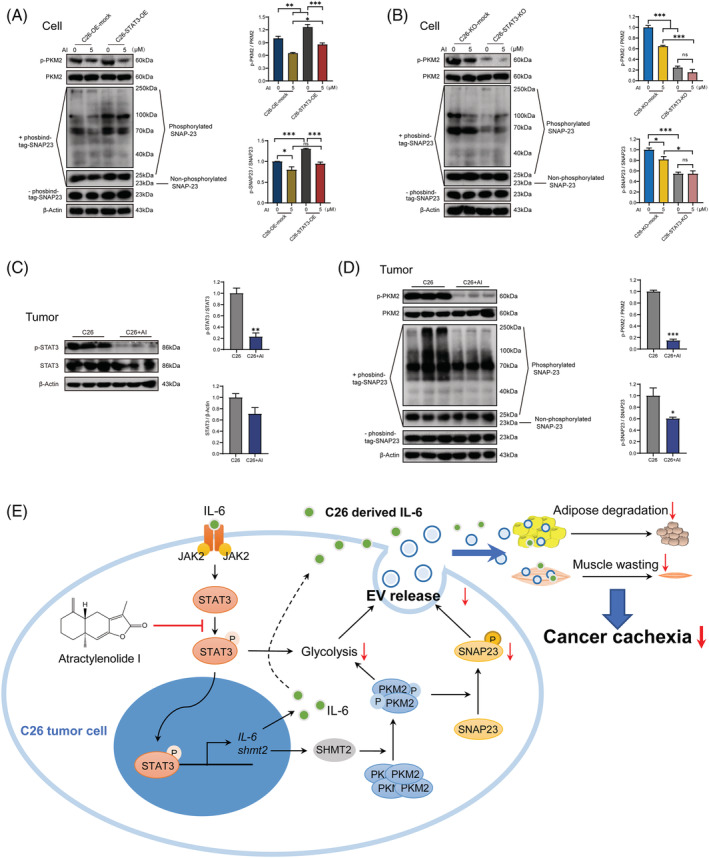
Methods: C26 tumour-bearing BALB/c mice were applied as animal model to examine the effects of AI (25 mg/kg) in attenuating cachexia symptoms, serum IL-6 and EVs levels. IL-6 and EVs secretion of C26 tumour cells treated with AI (0.31-5 μM) was further observed in vitro. The in vitro cultured C2C12 myotubes and 3T3-L1 mature adipocytes were used to check the potency of conditioned medium of C26 cells treated with AI (0.625-5 μM) in inducing muscle atrophy and lipolysis. The glycolysis potency of C26 cells under AI (0.31-5 μM) treatment was evaluated by measuring the extracellular acidification rate using Seahorse XFe96 Analyser. Levels of related signal proteins in both in vitro and in vivo experiments were examined using western blotting to study the possible mechanisms. STAT3 overexpression or knockout C26 cells were also used to confirm the effects of AI (5 μM).
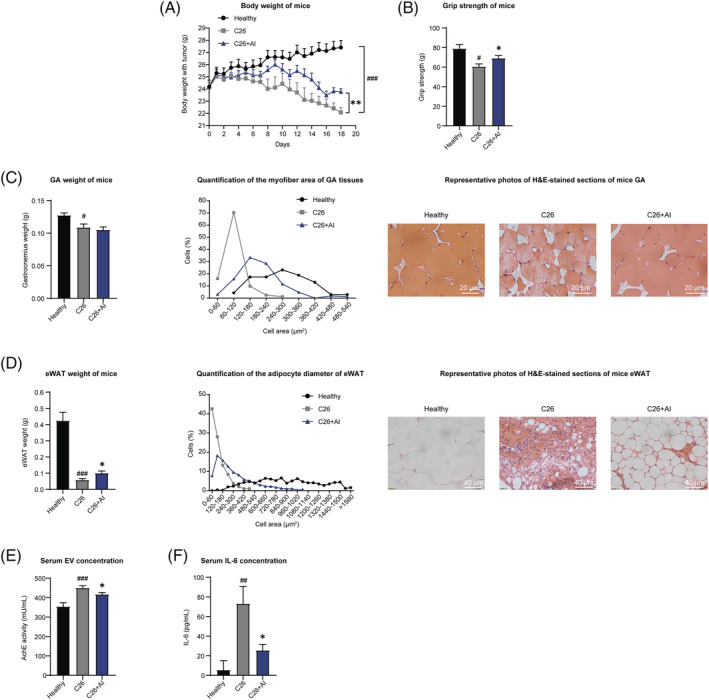
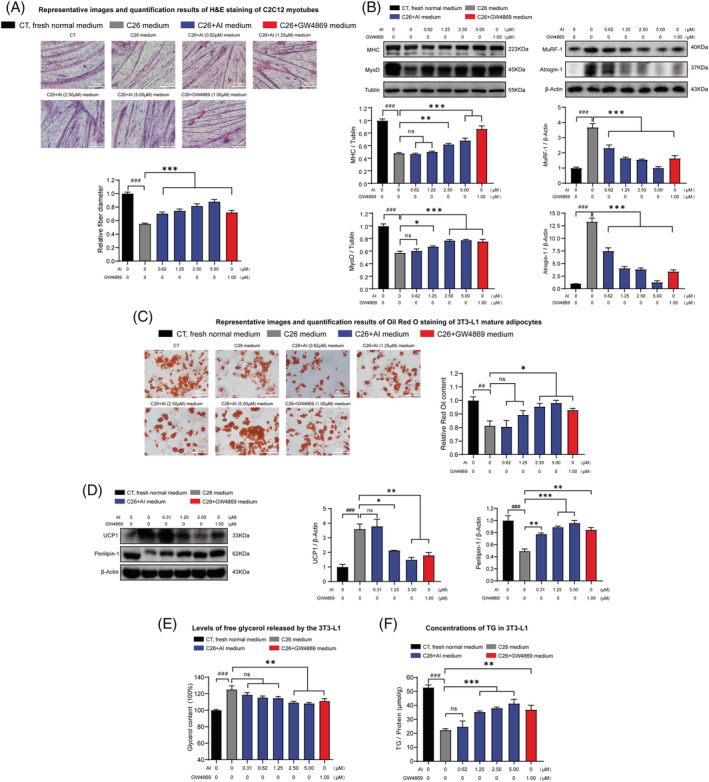
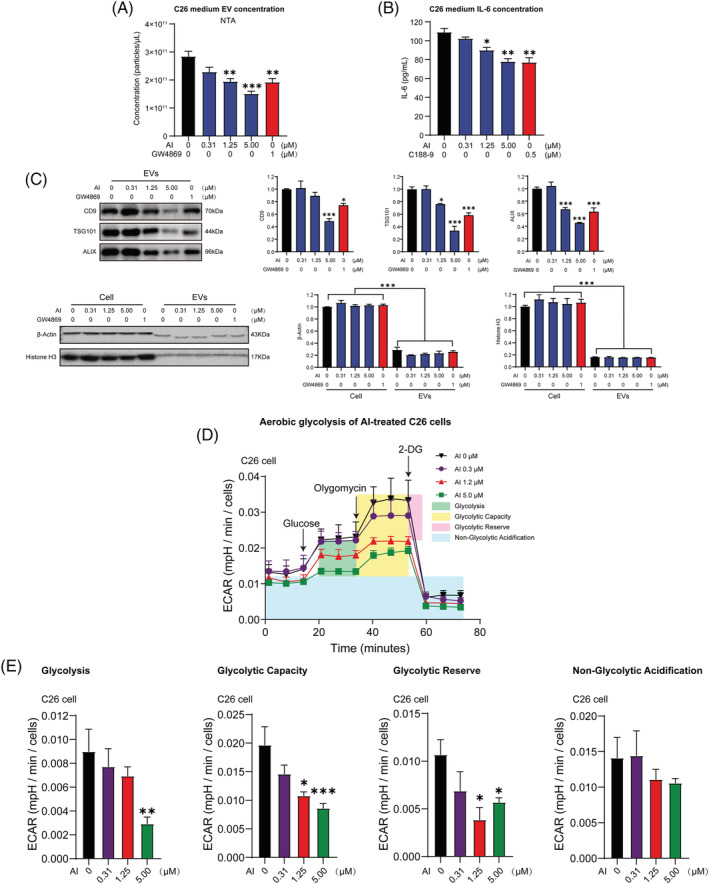
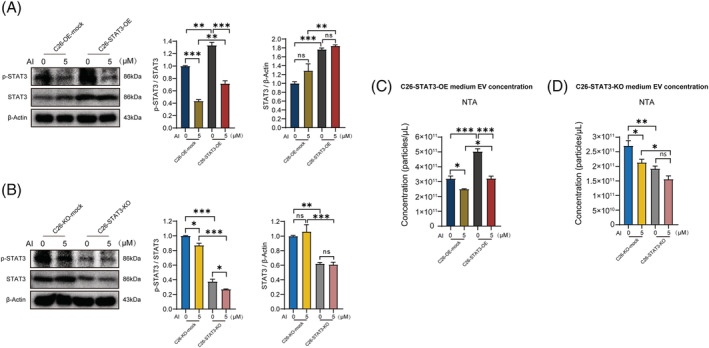
Results: AI ameliorated cancer cachexia symptoms (P < 0.05), improved grip strength (P < 0.05) and decreased serum EVs (P < 0.05) and IL-6 (P < 0.05) levels of C26 tumour-bearing mice. AI directly inhibited EVs biogenesis (P < 0.001) and IL-6 secretion (P < 0.01) of cultured C26 cells. The potency of C26 medium in inducing C2C12 myotube atrophy (+59.54%, P < 0.001) and 3T3-L1 adipocyte lipolysis (+20.73%, P < 0.05) was significantly attenuated when C26 cells were treated with AI. AI treatment inhibited aerobic glycolysis and the pathway of STAT3/PKM2/SNAP23 in C26 cells. Furthermore, overexpression of STAT3 partly antagonized the effects of AI in suppressing STAT3/PKM2/SNAP23 pathway, EVs secretion, glycolysis and the potency of C26 medium in inducing muscle atrophy and lipolysis, whereas knockout of STAT3 enhanced the inhibitory effect of AI on these values. The inhibition of AI on STAT3/PKM2/SNAP23 pathway was also observed in C26 tumour tissues.
Conclusions: AI ameliorates cancer cachexia by decreasing the production of IL-6 and EVs of tumour cells. The decreasing effects of AI on EVs biogenesis are based on its inhibition on STAT3/PKM2/SNAP23 pathway.
Keywords: STAT3; atractylenolide I; cancer cachexia; extracellular vesicle; lipolysis; muscle atrophy.
© 2022 The Authors. Journal of Cachexia, Sarcopenia and Muscle published by John Wiley & Sons Ltd on behalf of Society on Sarcopenia, Cachexia and Wasting Disorders.
Conflict of interest statement
All authors declare that they have no conflict of interest.
Figures
Similar articles
-
Lung cancer-derived extracellular vesicles induced myotube atrophy and adipocyte lipolysis via the extracellular IL-6-mediated STAT3 pathway.Biochim Biophys Acta Mol Cell Biol Lipids. 2019 Aug;1864(8):1091-1102. doi: 10.1016/j.bbalip.2019.04.006. Epub 2019 Apr 17. Biochim Biophys Acta Mol Cell Biol Lipids. 2019. PMID: 31002945
-
Alantolactone ameliorates cancer cachexia-associated muscle atrophy mainly by inhibiting the STAT3 signaling pathway.Phytomedicine. 2022 Jan;95:153858. doi: 10.1016/j.phymed.2021.153858. Epub 2021 Nov 15. Phytomedicine. 2022. PMID: 34861585
-
Carnosol and its analogues attenuate muscle atrophy and fat lipolysis induced by cancer cachexia.J Cachexia Sarcopenia Muscle. 2021 Jun;12(3):779-795. doi: 10.1002/jcsm.12710. Epub 2021 May 5. J Cachexia Sarcopenia Muscle. 2021. PMID: 33951335 Free PMC article.
-
Unravelling the Role of Cancer Cell-Derived Extracellular Vesicles in Muscle Atrophy, Lipolysis, and Cancer-Associated Cachexia.Cells. 2023 Nov 9;12(22):2598. doi: 10.3390/cells12222598. Cells. 2023. PMID: 37998333 Free PMC article. Review.
-
Extracellular Vesicles for Muscle Atrophy Treatment.Adv Exp Med Biol. 2023;1418:119-126. doi: 10.1007/978-981-99-1443-2_8. Adv Exp Med Biol. 2023. PMID: 37603276 Review.
Cited by
-
Screening of pivotal oncogenes modulated by DNA methylation in hepatocellular carcinoma and identification of atractylenolide I as an anti-cancer drug.Hum Cell. 2025 May 5;38(4):97. doi: 10.1007/s13577-025-01224-9. Hum Cell. 2025. PMID: 40325252
-
Race-based differences in serum biomarkers for cancer-associated cachexia in a diverse cohort of patients with pancreatic ductal adenocarcinoma.Res Sq [Preprint]. 2025 Feb 10:rs.3.rs-5690506. doi: 10.21203/rs.3.rs-5690506/v1. Res Sq. 2025. PMID: 39989973 Free PMC article. Preprint.
-
STAT3 Signalling Drives LDH Up-Regulation and Adiponectin Down-Regulation in Cachectic Adipocytes.Int J Mol Sci. 2023 Nov 15;24(22):16343. doi: 10.3390/ijms242216343. Int J Mol Sci. 2023. PMID: 38003534 Free PMC article.
-
Carnosol ameliorated cancer cachexia-associated myotube atrophy by targeting P5CS and its downstream pathways.Front Pharmacol. 2024 Jan 5;14:1291194. doi: 10.3389/fphar.2023.1291194. eCollection 2023. Front Pharmacol. 2024. PMID: 38249348 Free PMC article.
-
Potential Therapeutic Strategies for Skeletal Muscle Atrophy.Antioxidants (Basel). 2022 Dec 26;12(1):44. doi: 10.3390/antiox12010044. Antioxidants (Basel). 2022. PMID: 36670909 Free PMC article. Review.
References
-
- Argilés JM, Busquets S, Stemmler B, López‐Soriano FJ. Cancer cachexia: understanding the molecular basis. Nat Rev Cancer 2014;14:754–762. - PubMed
-
- Bailly C. Atractylenolides, essential components of Atractylodes‐based traditional herbal medicines: antioxidant, anti‐inflammatory and anticancer properties. Eur J Pharmacol 2021;891:173735. - PubMed
-
- Xiaomei R, Wenwen Q, Bao X. Effects of modified Sijunzi decoction and nutrition preparation on nutritional state of cancer patients. West J Trad Chin Med 2021;34:81–83.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
