

Therapeutic high affinity T cell receptor targeting a KRASG12D cancer neoantigen
- PMID: 36088370
- PMCID: PMC9464187
- DOI: 10.1038/s41467-022-32811-1
Therapeutic high affinity T cell receptor targeting a KRASG12D cancer neoantigen
Abstract
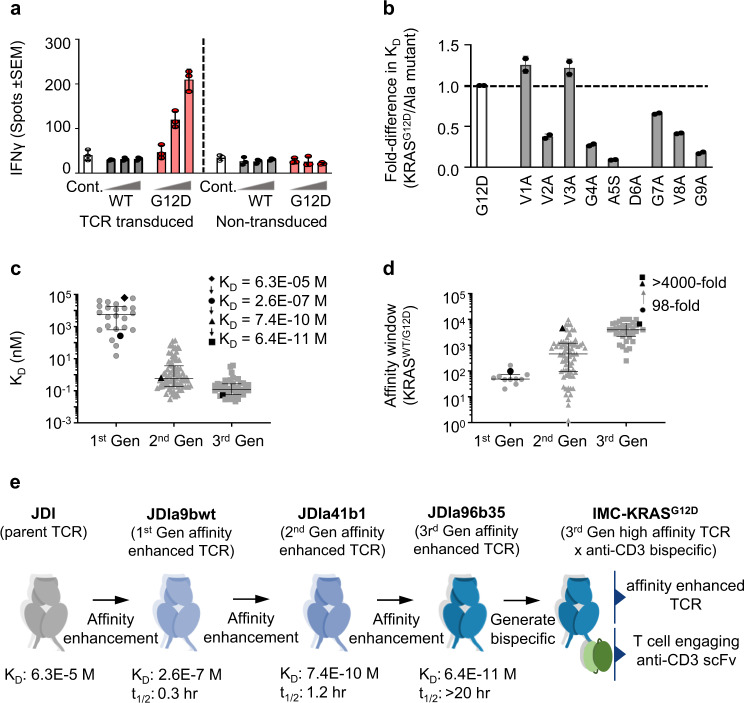
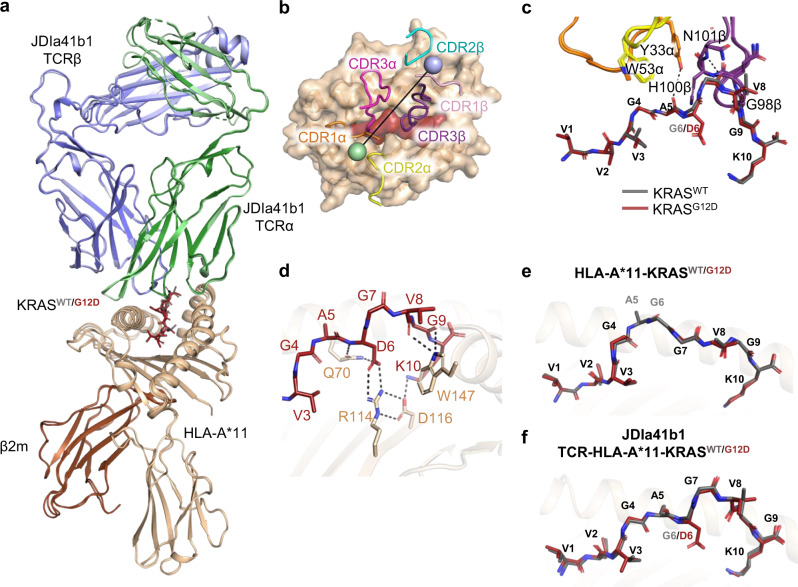
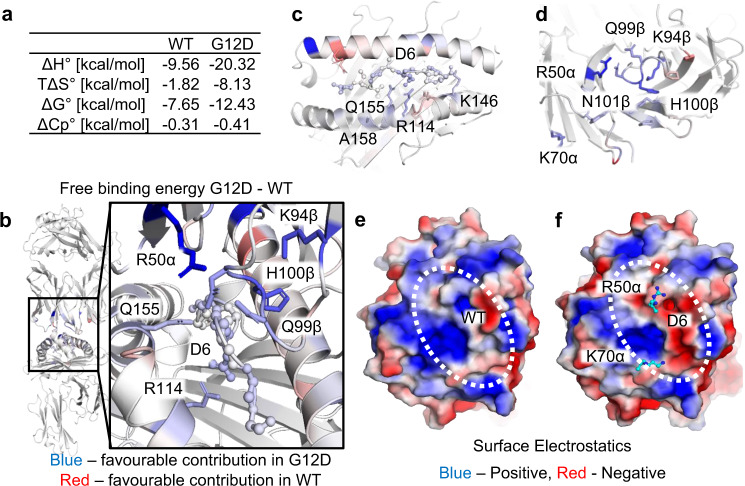
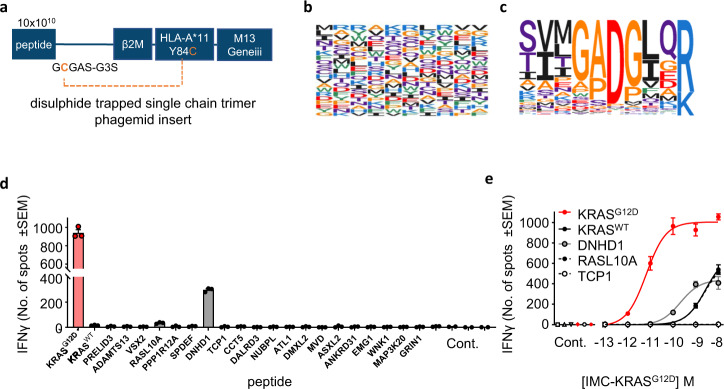
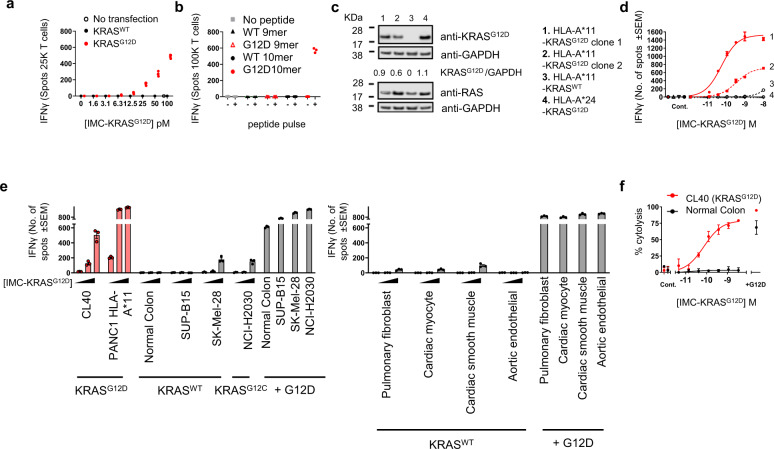
Neoantigens derived from somatic mutations are specific to cancer cells and are ideal targets for cancer immunotherapy. KRAS is the most frequently mutated oncogene and drives the pathogenesis of several cancers. Here we show the identification and development of an affinity-enhanced T cell receptor (TCR) that recognizes a peptide derived from the most common KRAS mutant, KRASG12D, presented in the context of HLA-A*11:01. The affinity of the engineered TCR is increased by over one million-fold yet fully able to distinguish KRASG12D over KRASWT. While crystal structures reveal few discernible differences in TCR interactions with KRASWT versus KRASG12D, thermodynamic analysis and molecular dynamics simulations reveal that TCR specificity is driven by differences in indirect electrostatic interactions. The affinity enhanced TCR, fused to a humanized anti-CD3 scFv, enables selective killing of cancer cells expressing KRASG12D. Our work thus reveals a molecular mechanism that drives TCR selectivity and describes a soluble bispecific molecule with therapeutic potential against cancers harboring a common shared neoantigen.
© 2022. The Author(s).
Conflict of interest statement
A.P., V.K., S.M., T.D., C.H., C.R., J.D., S.H., K.B., M.H., C.C., M.A., R.R., J.D.D., N.L., A.V., D.K.C., A.D.W., and C.C. are current or former employees of, and may hold shares in Immunocore Ltd. J.N.H, W.Y., A.B.L., and G.D.P. are current or former employees of, and may hold shares in Eli Lilly and Co. M.v.d.K and A.H. declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
