

The PCI domains are "winged" HEAT domains
- PMID: 36094910
- PMCID: PMC9467303
- DOI: 10.1371/journal.pone.0268664
The PCI domains are "winged" HEAT domains
Abstract
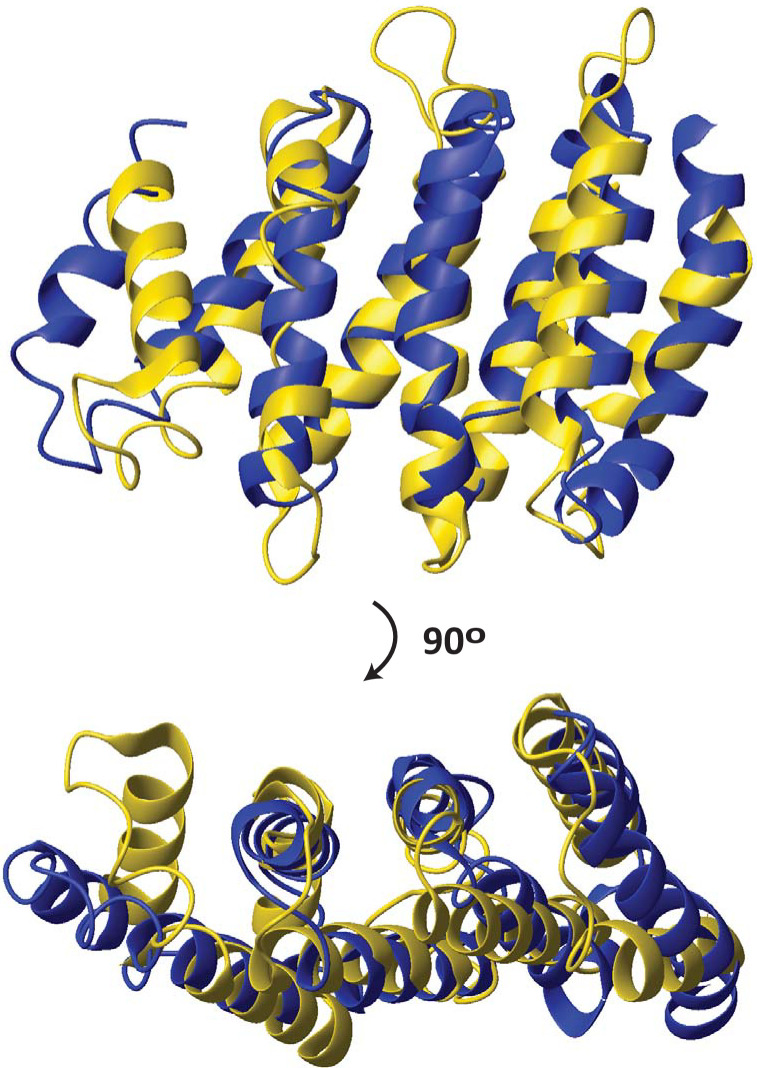
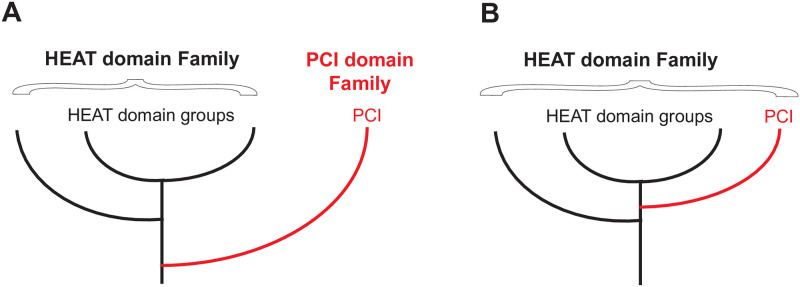
The HEAT domains are a family of helical hairpin repeat domains, composed of four or more hairpins. HEAT is derived from the names of four family members: huntingtin, eukaryotic translation elongation factor 3 (eEF3), protein phosphatase 2 regulatory A subunit (PP2A), and mechanistic target of rapamycin (mTOR). HEAT domain-containing proteins play roles in a wide range of cellular processes, such as protein synthesis, nuclear transport and metabolism, and cell signaling. The PCI domains are a related group of helical hairpin domains, with a "winged-helix" (WH) subdomain at their C-terminus, which is responsible for multi-subunit complex formation with other PCI domains. The name is derived from the complexes, where these domains are found: the 26S Proteasome "lid" regulatory subcomplex, the COP9 signalosome (CSN), and eukaryotic translation initiation factor 3 (eIF3). We noted that in structure similarity searches using HEAT domains, sometimes PCI domains appeared in the search results ahead of other HEAT domains, which indicated that the PCI domains could be members of the HEAT domain family, and not a related but separate group, as currently thought. Here, we report extensive structure similarity analysis of HEAT and PCI domains, both within and between the two groups of proteins. We present evidence that the PCI domains as a group have greater structural similarity with individual groups of HEAT domains than some of the HEAT domain groups have among each other. Therefore, our results indicate that the PCI domains have evolved from a HEAT domain that acquired a WH subdomain. The WH subdomain in turn mediated self-association into a multi-subunit complex, which eventually evolved into the common ancestor of the Proteasome lid/CSN/eIF3.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures



References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
