

Transmembrane helices mediate the formation of a stable ternary complex of b5R, cyt b5, and SCD1
- PMID: 36097052
- PMCID: PMC9468158
- DOI: 10.1038/s42003-022-03882-z
Transmembrane helices mediate the formation of a stable ternary complex of b5R, cyt b5, and SCD1
Abstract
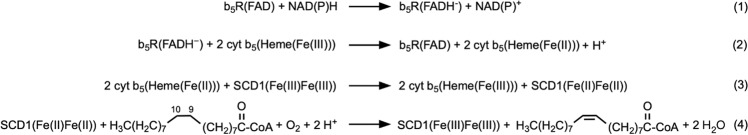
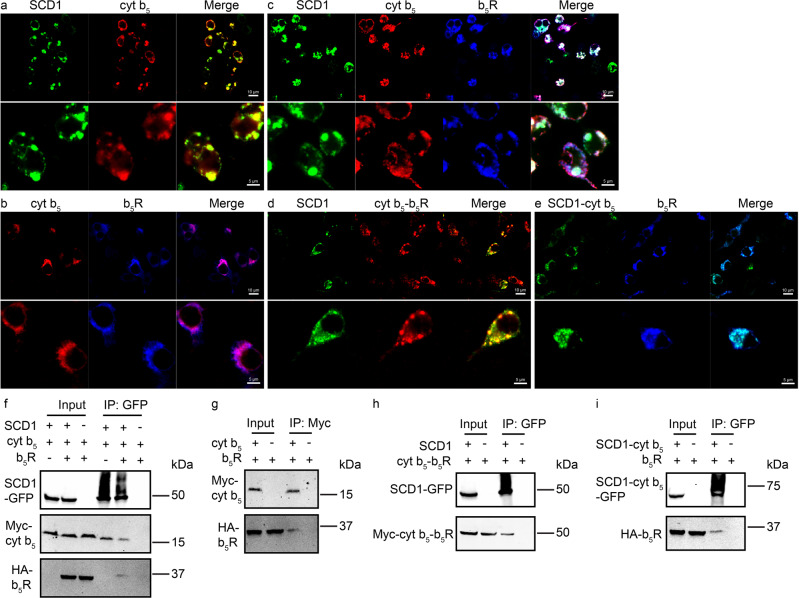
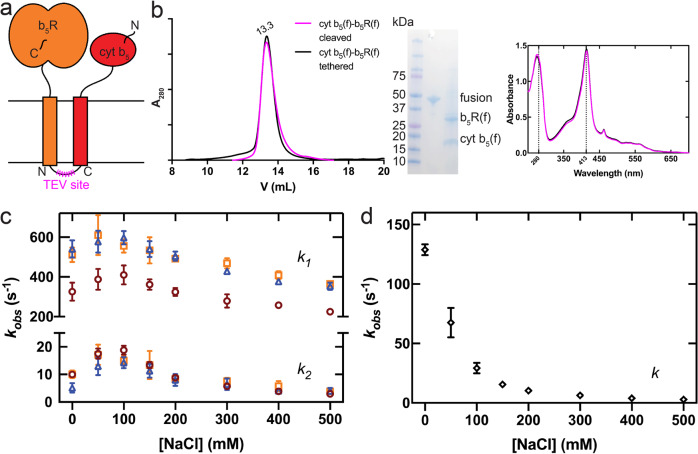
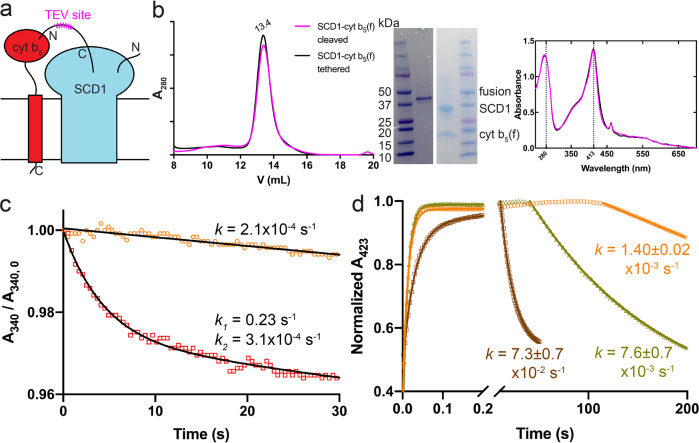
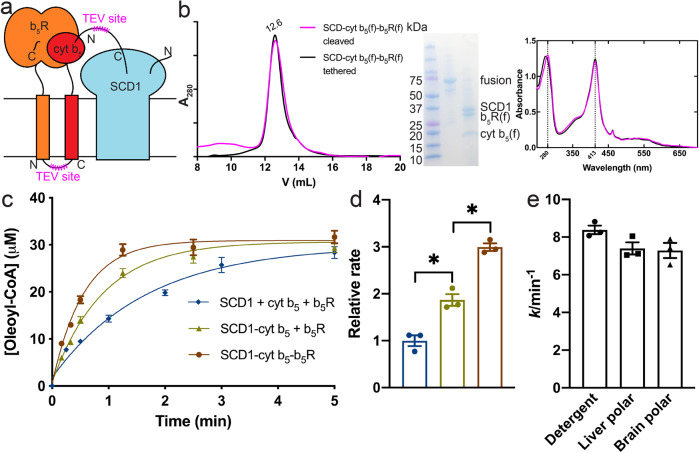
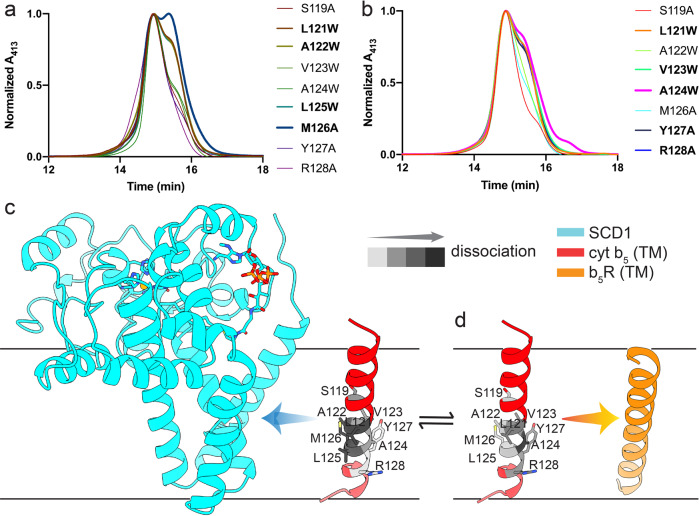
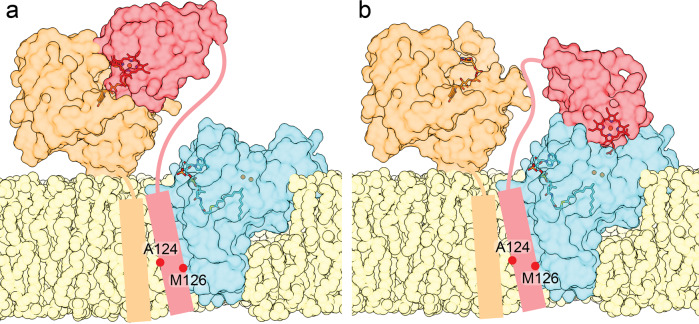
Mammalian cytochrome b5 (cyt b5) and cytochrome b5 reductase (b5R) are electron carrier proteins for membrane-embedded oxidoreductases. Both b5R and cyt b5 have a cytosolic domain and a single transmembrane (TM) helix. The cytosolic domains of b5R and cyt b5 contain cofactors required for electron transfer, but it is not clear if the TM helix has function beyond being an anchor to the membrane. Here we show that b5R and cyt b5 form a stable binary complex, and so do cyt b5 and stearoyl-CoA desaturase-1 (SCD1). We also show that b5R, cyt b5 and SCD1 form a stable ternary complex. We demonstrate that the TM helices are required for the assembly of stable binary and ternary complexes where electron transfer rates are greatly enhanced. These results reveal a role of the TM helix in cyt b5 and b5R, and suggest that an electron transport chain composed of a stable ternary complex may be a general feature in membrane-embedded oxidoreductases that require cyt b5 and b5R.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
