

The Single-Cell Immunogenomic Landscape of B and Plasma Cells in Early-Stage Lung Adenocarcinoma
- PMID: 36098652
- PMCID: PMC9633381
- DOI: 10.1158/2159-8290.CD-21-1658
The Single-Cell Immunogenomic Landscape of B and Plasma Cells in Early-Stage Lung Adenocarcinoma
Abstract
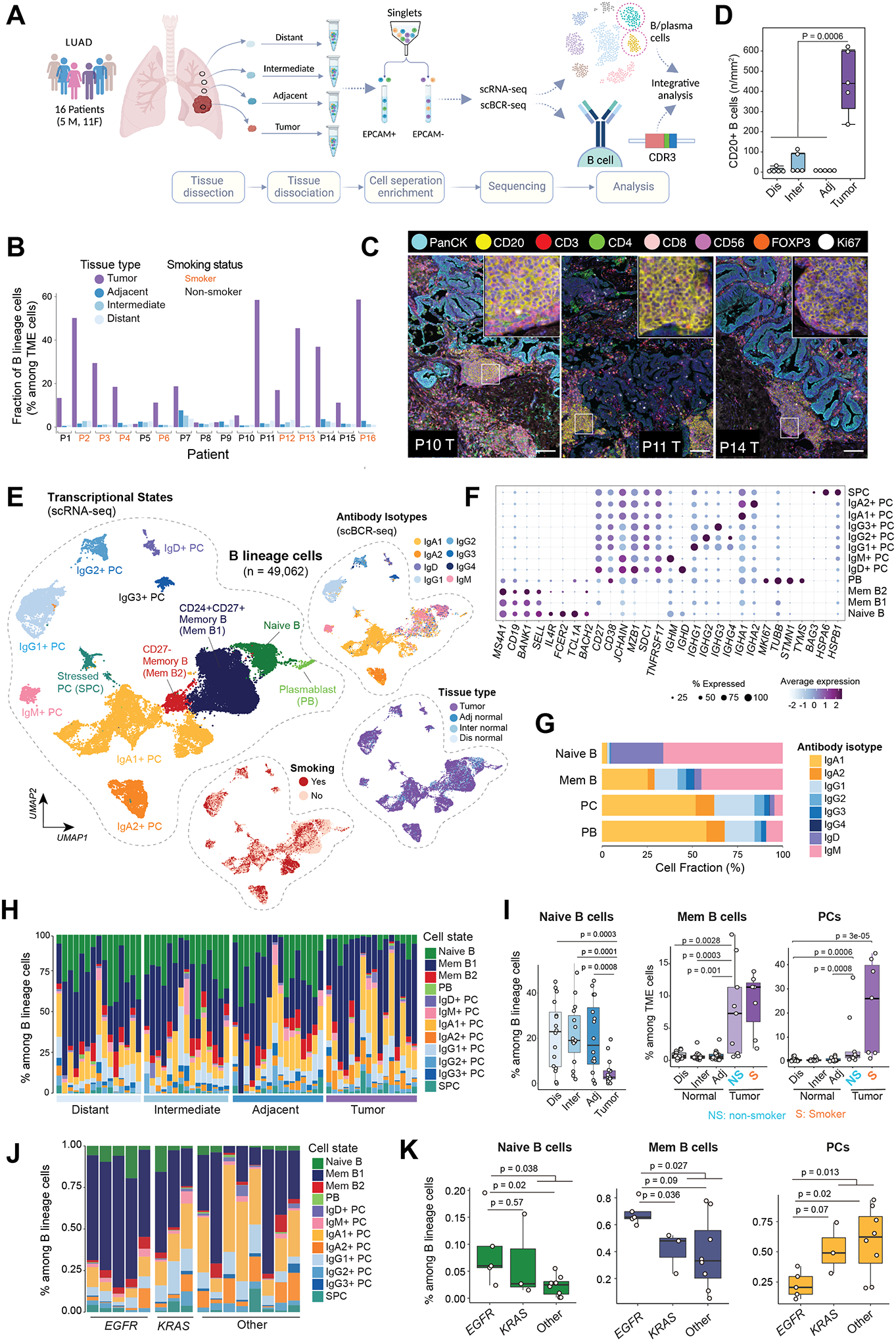
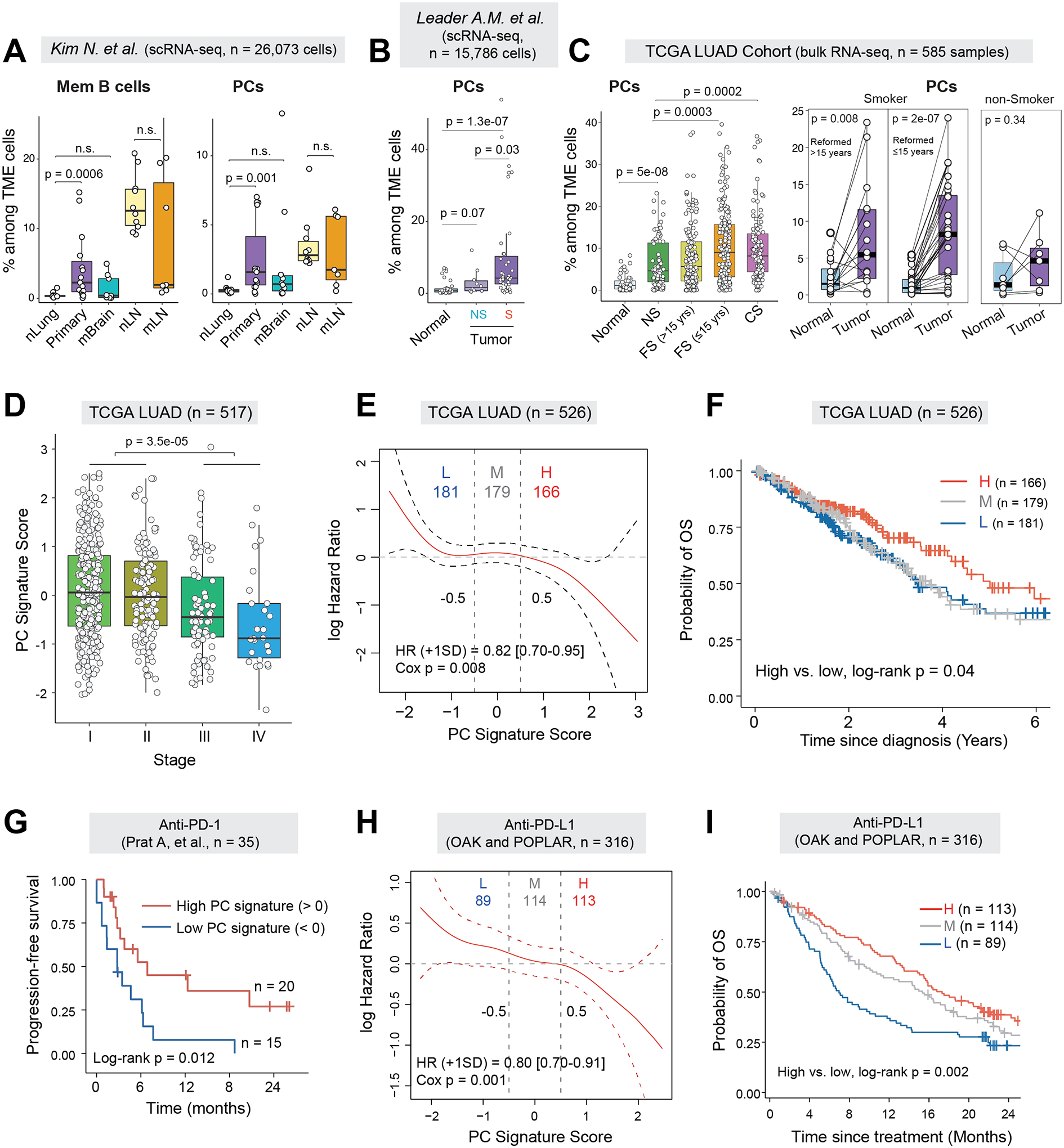
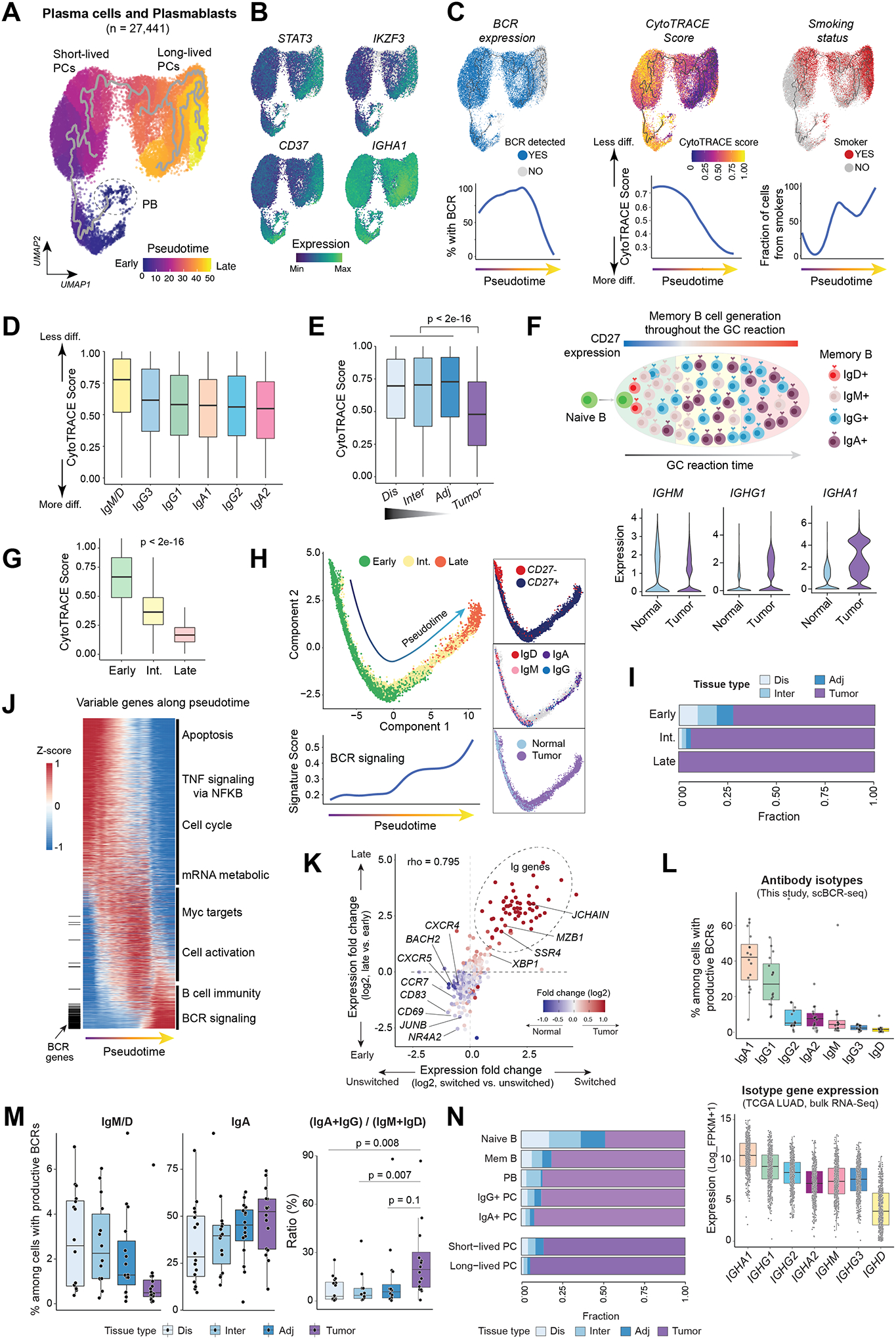
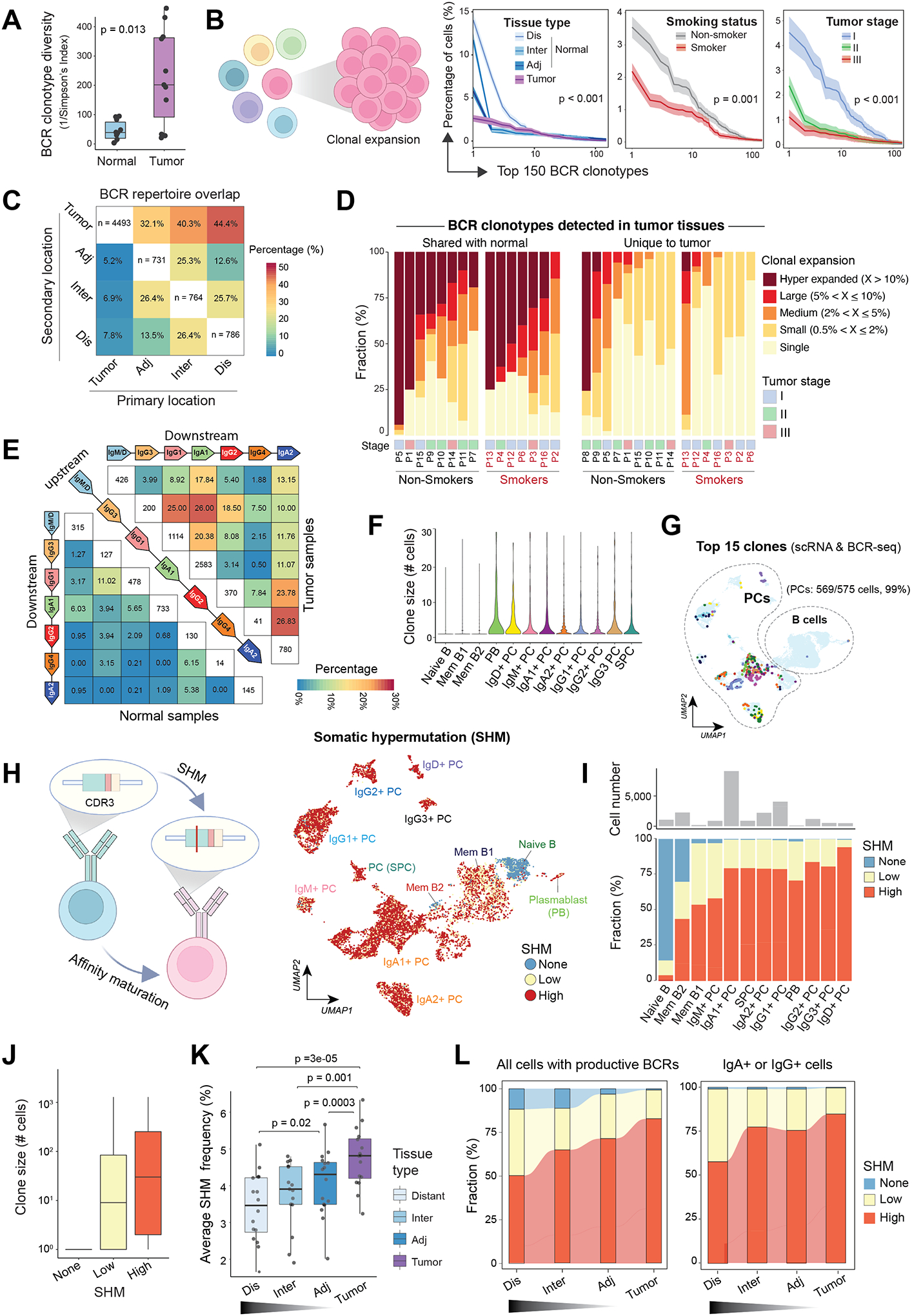
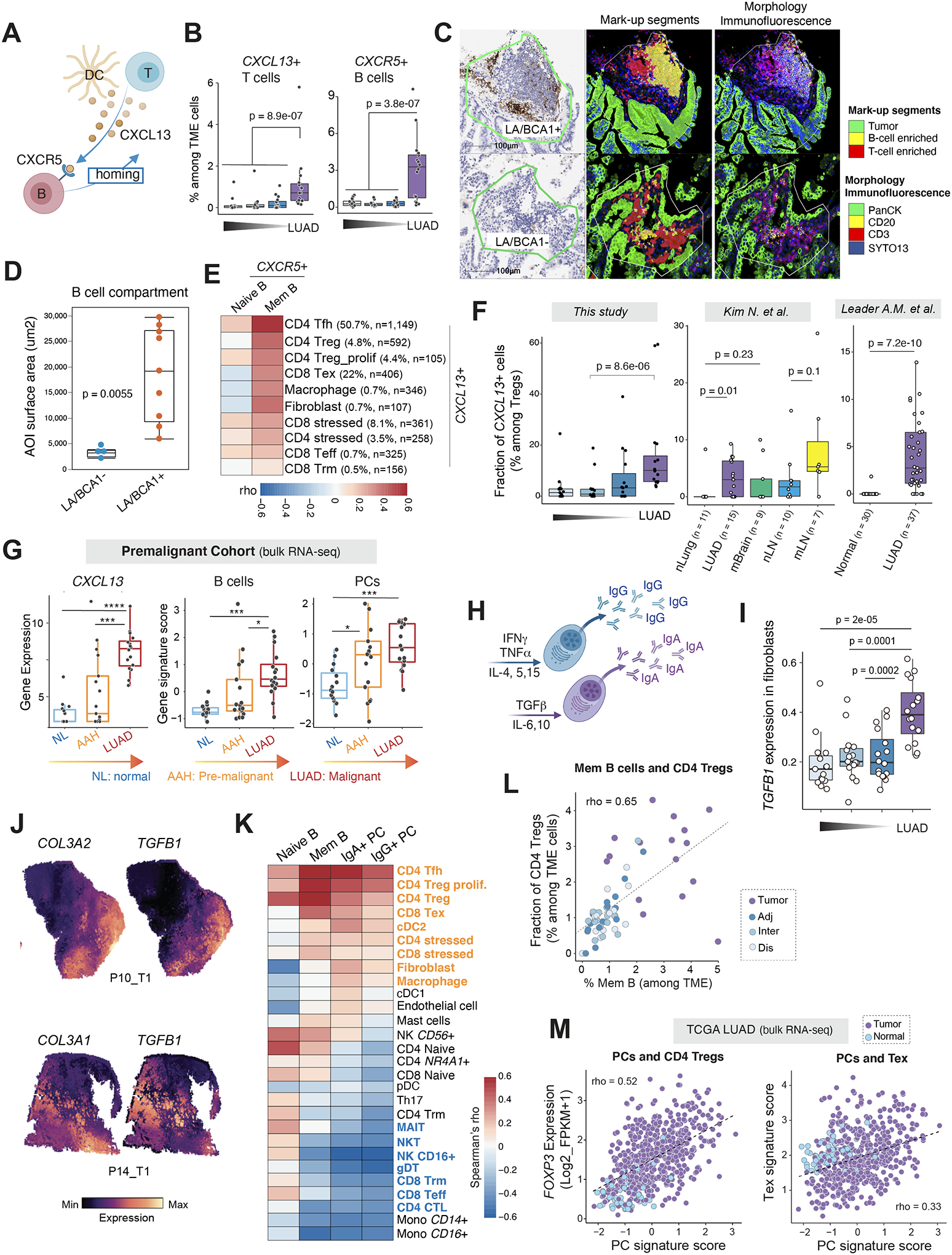
Tumor-infiltrating B and plasma cells (TIB) are prevalent in lung adenocarcinoma (LUAD); however, they are poorly characterized. We performed paired single-cell RNA and B-cell receptor (BCR) sequencing of 16 early-stage LUADs and 47 matching multiregion normal tissues. By integrative analysis of ∼50,000 TIBs, we define 12 TIB subsets in the LUAD and adjacent normal ecosystems and demonstrate extensive remodeling of TIBs in LUADs. Memory B cells and plasma cells (PC) were highly enriched in tumor tissues with more differentiated states and increased frequencies of somatic hypermutation. Smokers exhibited markedly elevated PCs and PCs with distinct differentiation trajectories. BCR clonotype diversity increased but clonality decreased in LUADs, smokers, and with increasing pathologic stage. TIBs were mostly localized within CXCL13+ lymphoid aggregates, and immune cell sources of CXCL13 production evolved with LUAD progression and included elevated fractions of CD4 regulatory T cells. This study provides a spatial landscape of TIBs in early-stage LUAD.
Significance: While TIBs are highly enriched in LUADs, they are poorly characterized. This study provides a much-needed understanding of the transcriptional, clonotypic states and phenotypes of TIBs, unraveling their potential roles in the immunopathology of early-stage LUADs and constituting a road map for the development of TIB-targeted immunotherapies for the treatment of this morbid malignancy. This article is highlighted in the In This Issue feature, p. 2483.
©2022 American Association for Cancer Research.
Figures

References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
