

Effect of Phosphate and Ferritin Subunit Composition on the Kinetics, Structure, and Reactivity of the Iron Core in Human Homo- and Heteropolymer Ferritins
- PMID: 36099002
- PMCID: PMC9548343
- DOI: 10.1021/acs.biochem.2c00354
Effect of Phosphate and Ferritin Subunit Composition on the Kinetics, Structure, and Reactivity of the Iron Core in Human Homo- and Heteropolymer Ferritins
Abstract
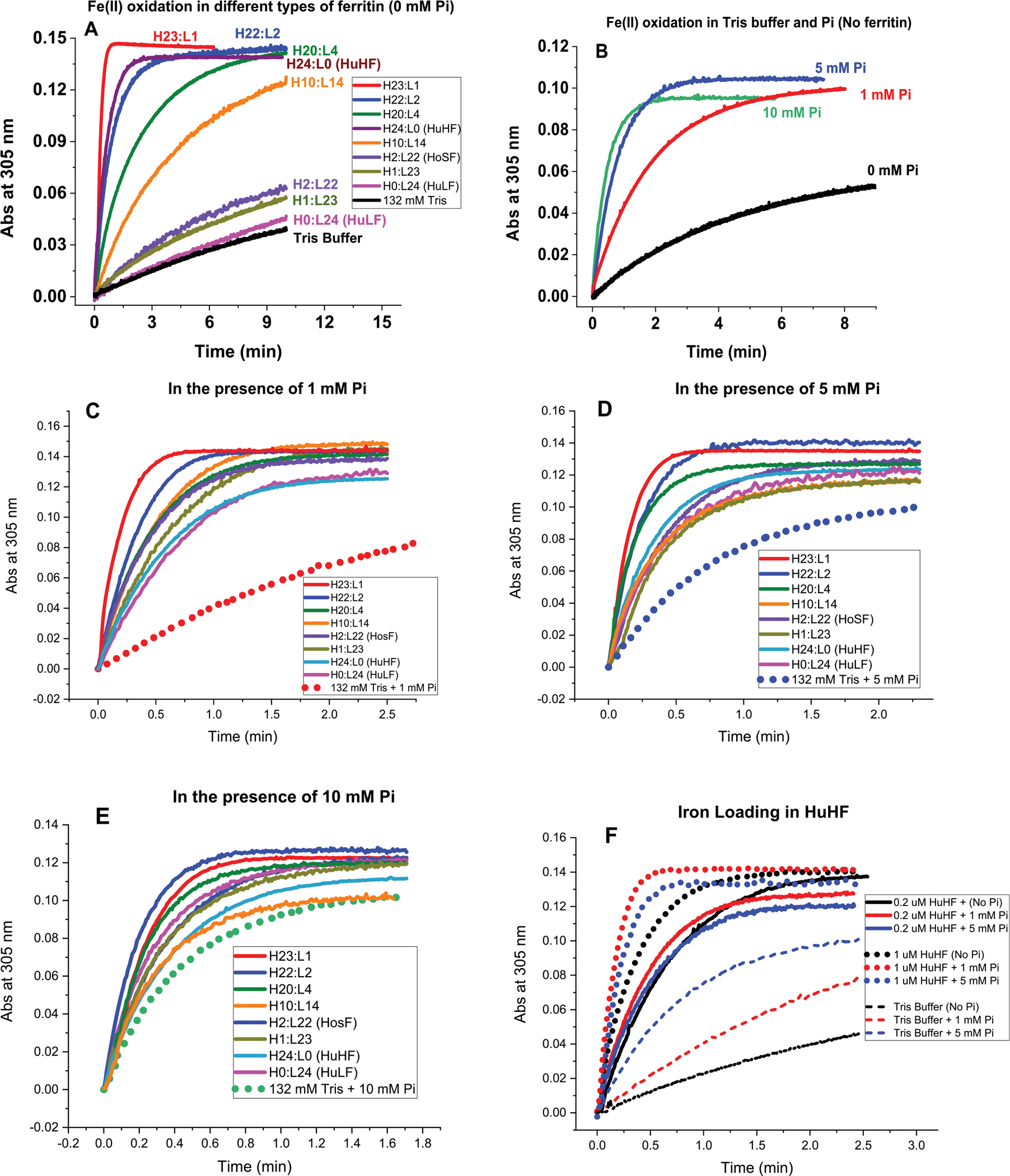
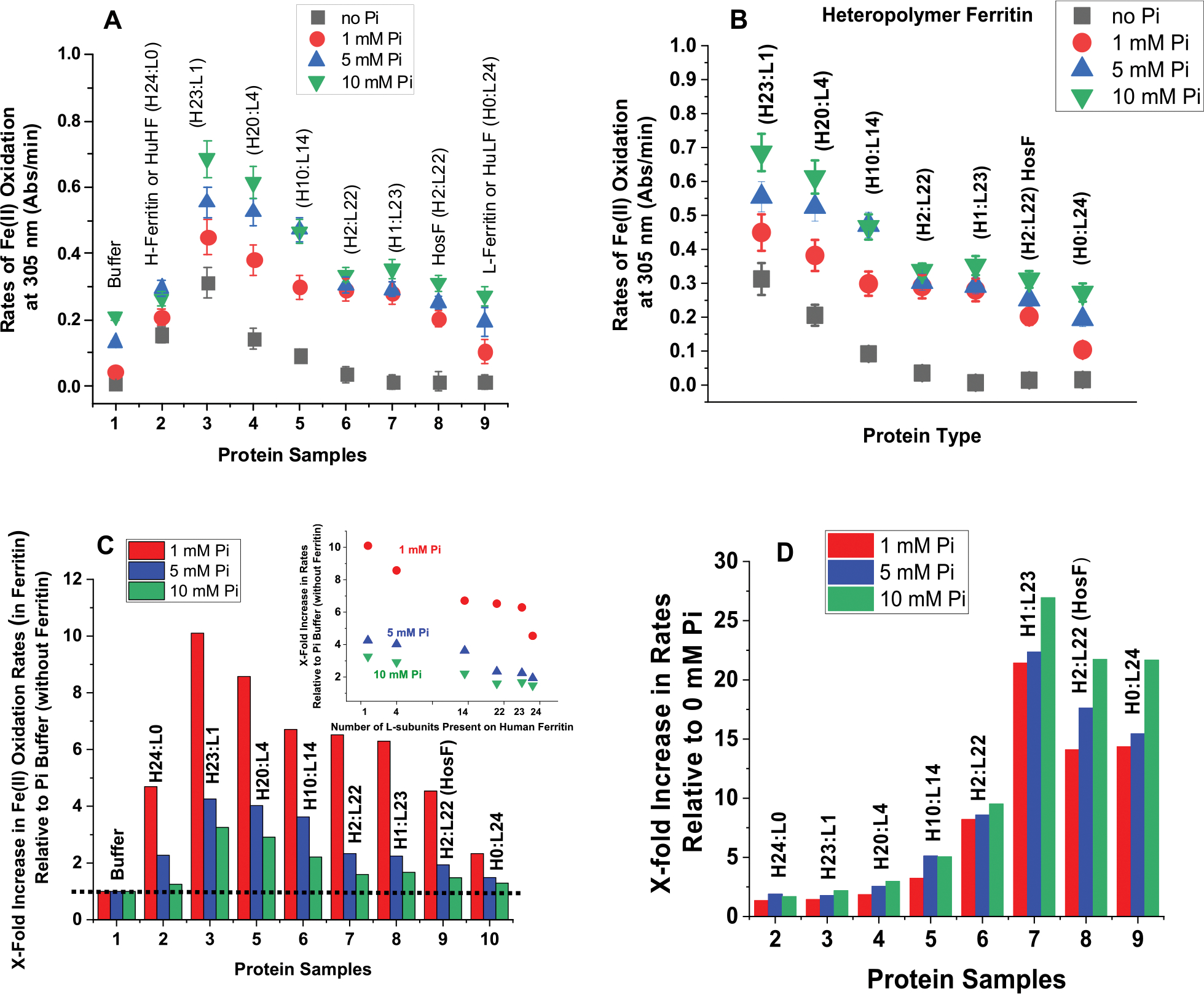
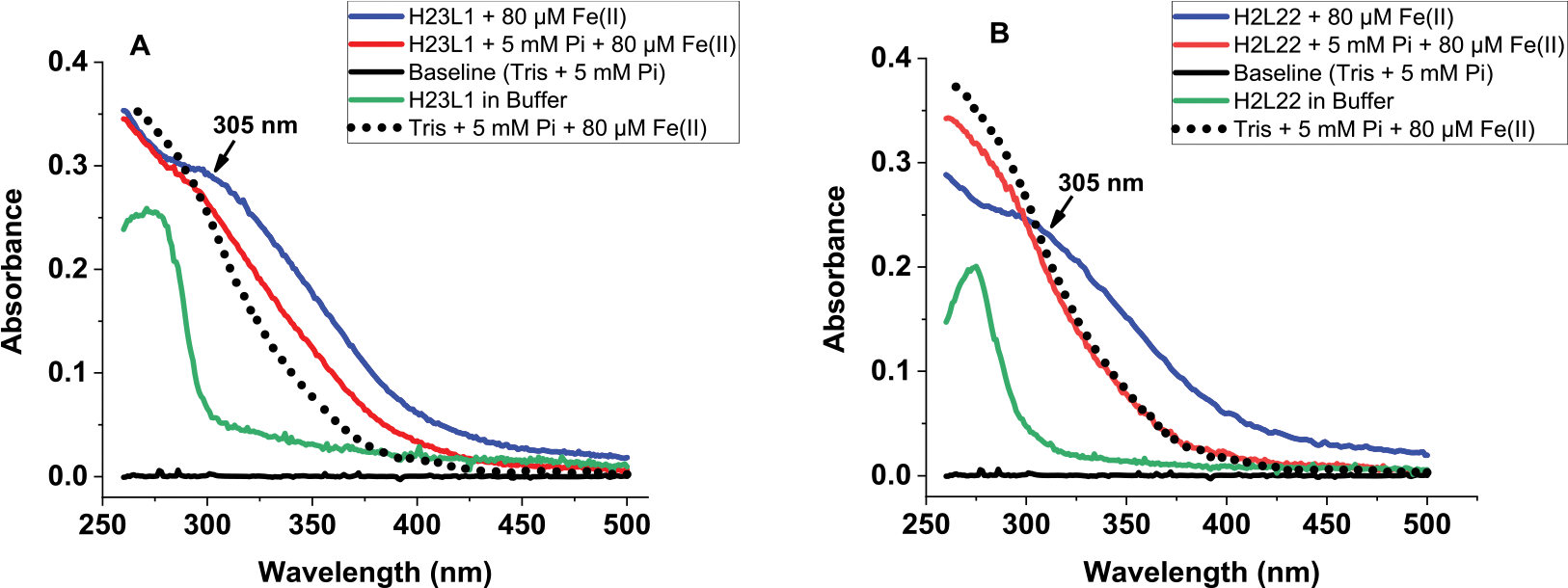
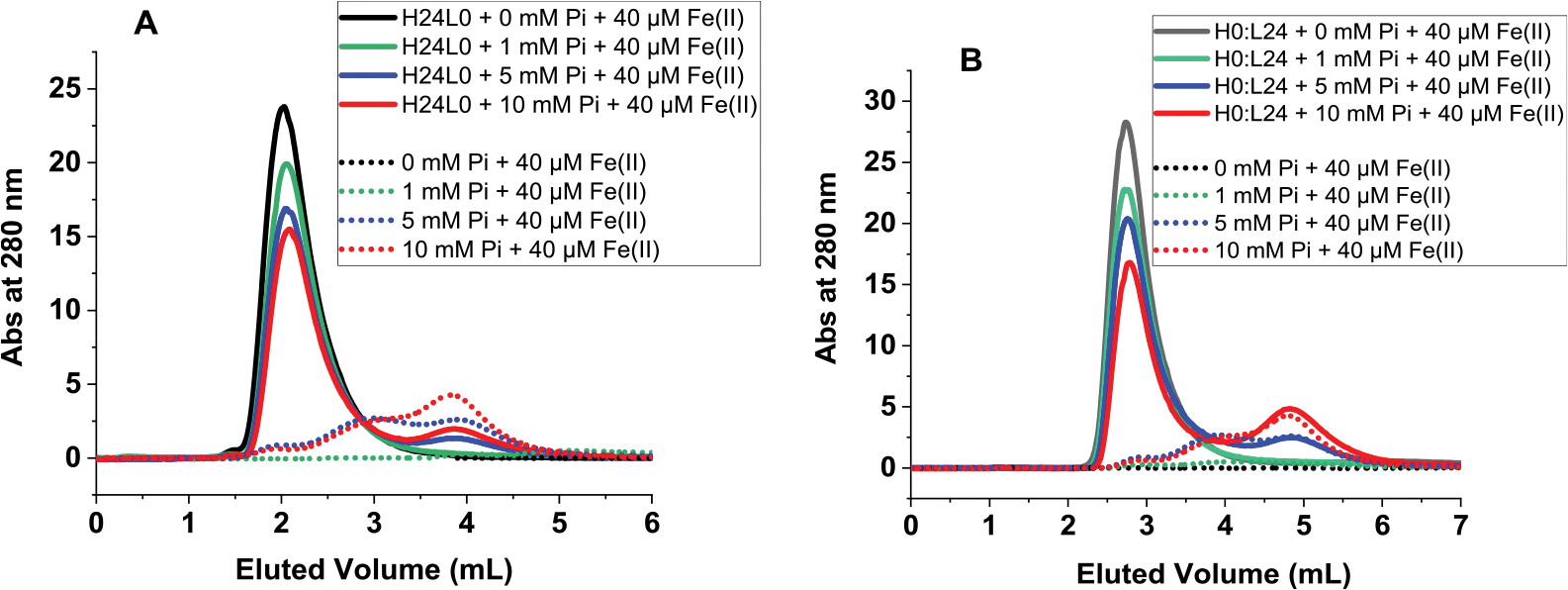
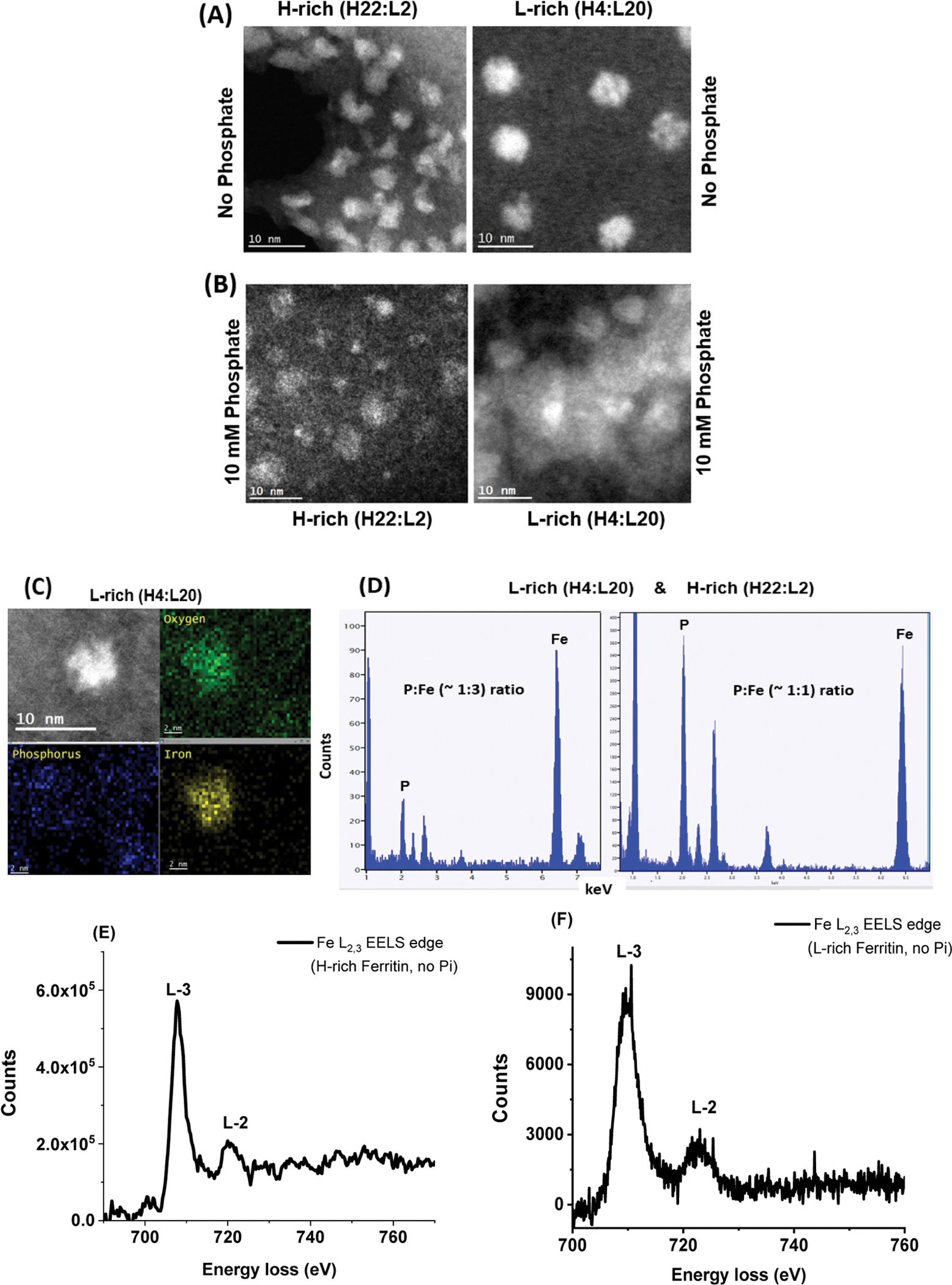
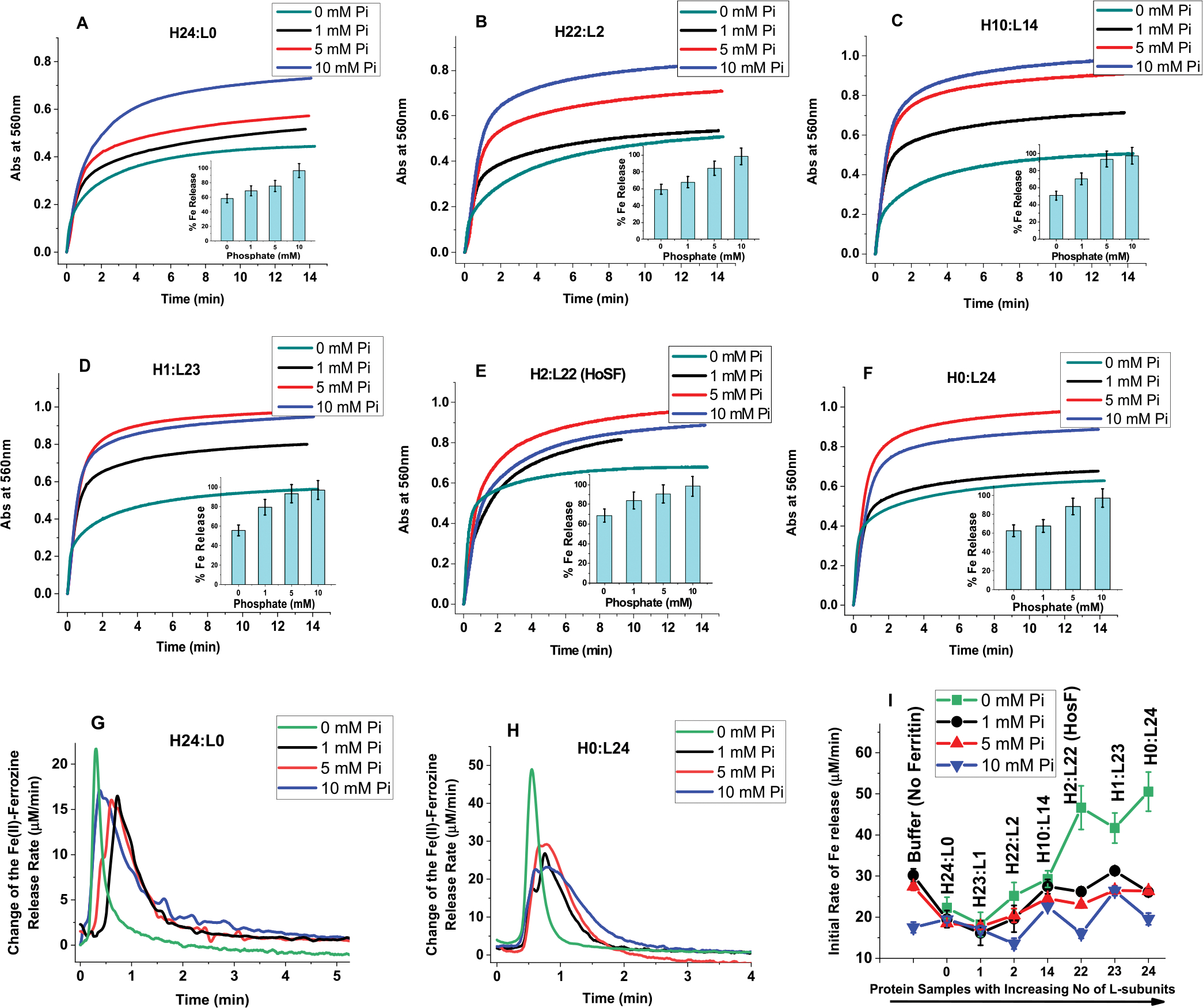
Ferritins are highly conserved supramolecular protein nanostructures that play a key role in iron homeostasis. Thousands of iron atoms can be stored inside their hollow cavity as a hydrated ferric oxyhydroxide mineral. Although phosphate associates with the ferritin iron nanoparticles, the effect of physiological concentrations on the kinetics, structure, and reactivity of ferritin iron cores has not yet been explored. Here, the iron loading and mobilization kinetics were studied in the presence of 1-10 mM phosphate using homopolymer and heteropolymer ferritins having different H to L subunit ratios. In the absence of ferritin, phosphate enhances the rate of ferrous ion oxidation and forms large and soluble polymeric Fe(III)-phosphate species. In the presence of phosphate, Fe(II) oxidation and core formation in ferritin is significantly accelerated with oxidation rates several-fold higher than with phosphate alone. High-angle annular dark-field scanning transmission electron microscopy measurements revealed a strong phosphate effect on both the size and morphology of the iron mineral in H-rich (but not L-rich) ferritins. While iron nanoparticles in L-rich ferritins have spherical shape in the absence and presence of phosphate, iron nanoparticles in H-rich ferritins change from irregular shapes in the absence of phosphate to spherical particles in the presence of phosphate with larger size distribution and smaller particle size. In the presence of phosphate, the kinetics of iron-reductive mobilization from ferritin releases twice as much iron than in its absence. Altogether, our results demonstrate an important role for phosphate, and the ferritin H and L subunit composition toward the kinetics of iron oxidation and removal from ferritin, as well as the structure and reactivity of the iron mineral, and may have an important implication on ferritin iron management in vivo.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
References
-
- Bou-Abdallah F The Iron Redox and Hydrolysis Chemistry of the Ferritins. Biochim. Biophys. Acta 2010, 1800, 719–731. - PubMed
-
- Ford GC; Harrison PM; Rice DW; Smith JM; Treffry A; White JL; Yariv J Ferritin: Design and Formation of an Iron-Storage Molecule. Philos. Trans. R. Soc. London, Ser. B 1984, 304, 551–565. - PubMed
-
- Harrison PM; Andrews SC; Artymiuk PJ; Ford GC; Guest JR; Hirzmann J; Lawson DM; Livingstone JC; Smith JMA; Treffry A; Yewdall SJ Probing Structure-Function Relations In Ferritin And Bacterioferritin. Adv. Inorg. Chem. 1991, 36, 449–486.
-
- Melman A; Bou-Abdallah F Iron Mineralization and Core Dissociation in Mammalian Homopolymeric H-Ferritin: Current Understanding and Future Perspectives. Biochim. Biophys. Acta 2020, 1864, 129700. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
