

Homologous recombination-deficient mutation cluster in tumor suppressor RAD51C identified by comprehensive analysis of cancer variants
- PMID: 36099300
- PMCID: PMC9499524
- DOI: 10.1073/pnas.2202727119
Homologous recombination-deficient mutation cluster in tumor suppressor RAD51C identified by comprehensive analysis of cancer variants
Abstract
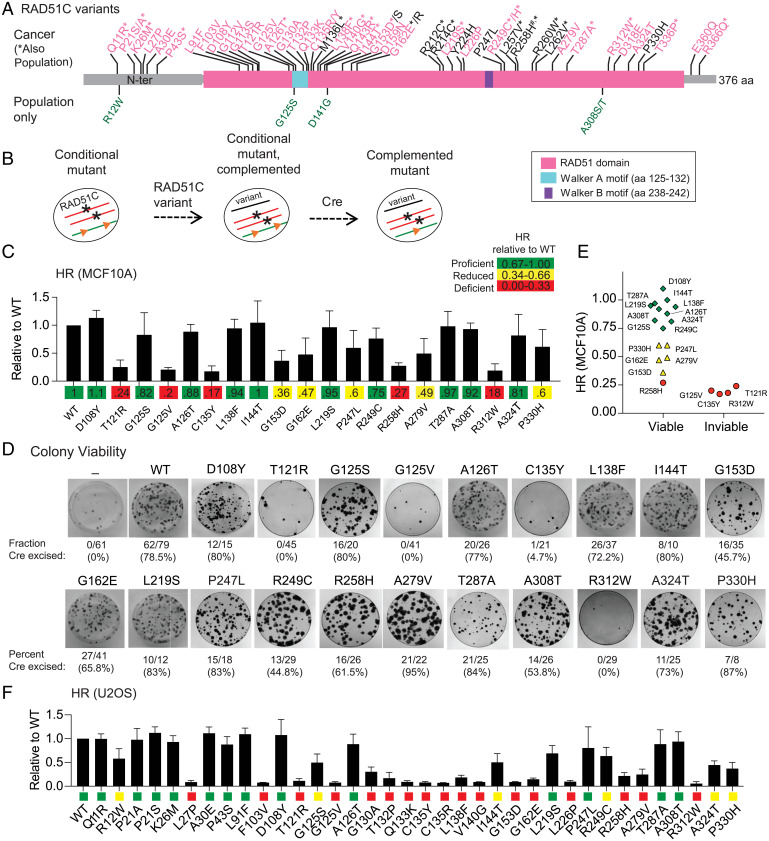
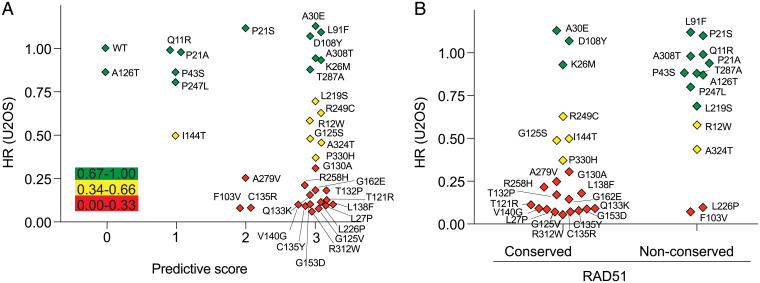
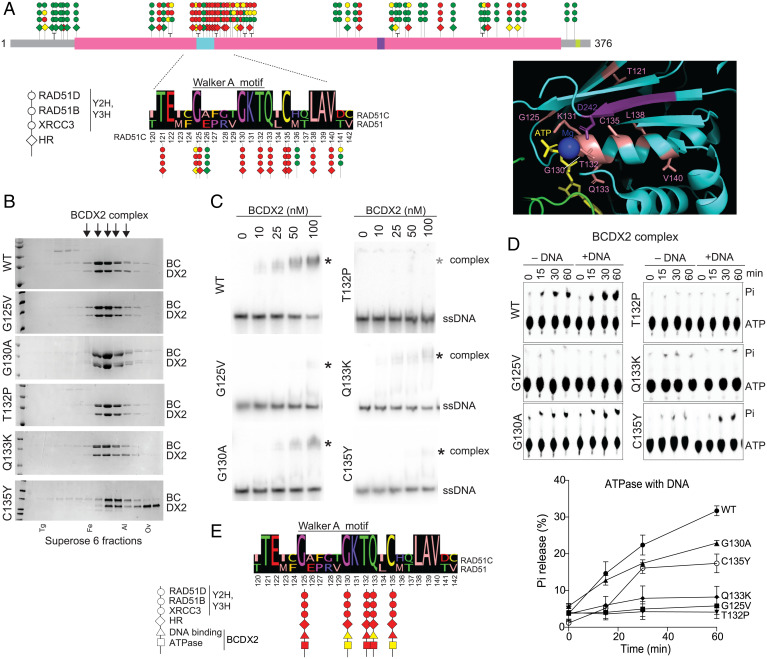
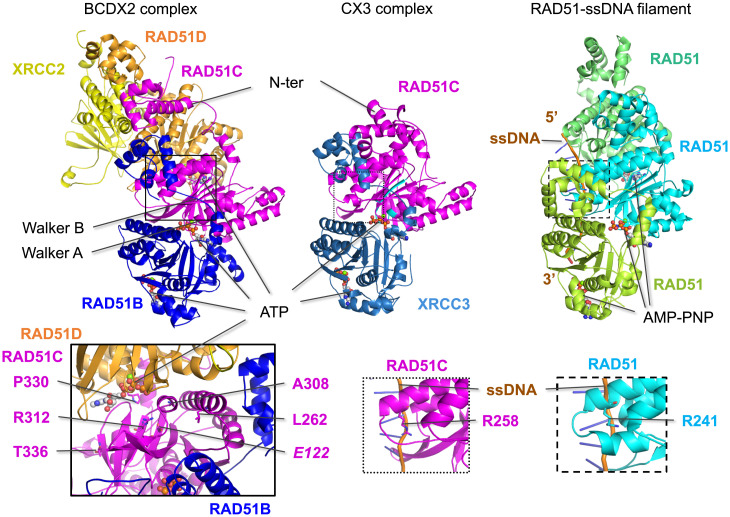
Mutations in homologous recombination (HR) genes, including BRCA1, BRCA2, and the RAD51 paralog RAD51C, predispose to tumorigenesis and sensitize cancers to DNA-damaging agents and poly(ADP ribose) polymerase inhibitors. However, ∼800 missense variants of unknown significance have been identified for RAD51C alone, impairing cancer risk assessment and therapeutic strategies. Here, we interrogated >50 RAD51C missense variants, finding that mutations in residues conserved with RAD51 strongly predicted HR deficiency and disrupted interactions with other RAD51 paralogs. A cluster of mutations was identified in and around the Walker A box that led to impairments in HR, interactions with three other RAD51 paralogs, binding to single-stranded DNA, and ATP hydrolysis. We generated structural models of the two RAD51 paralog complexes containing RAD51C, RAD51B-RAD51C-RAD51D-XRCC2 and RAD51C-XRCC3. Together with our functional and biochemical analyses, the structural models predict ATP binding at the interface of RAD51C interactions with other RAD51 paralogs, similar to interactions between monomers in RAD51 filaments, and explain the failure of RAD51C variants in binding multiple paralogs. Ovarian cancer patients with variants in this cluster showed exceptionally long survival, which may be relevant to the reversion potential of the variants. This comprehensive analysis provides a framework for RAD51C variant classification. Importantly, it also provides insight into the functioning of the RAD51 paralog complexes.
Keywords: DNA repair; Homologous recombination; RAD51 paralog; RAD51C; Variants of unknown significance.
Conflict of interest statement
The authors declare no competing interest.
Figures

References
-
- O’Connor M. J., Targeting the DNA damage response in cancer. Mol. Cell 60, 547–560 (2015). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
