

Perilipin-2 limits remyelination by preventing lipid droplet degradation
- PMID: 36100764
- PMCID: PMC11803036
- DOI: 10.1007/s00018-022-04547-0
Perilipin-2 limits remyelination by preventing lipid droplet degradation
Abstract
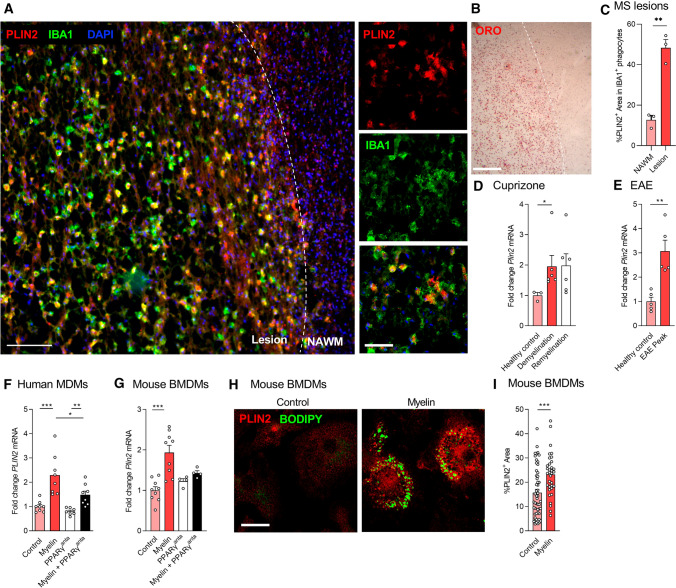
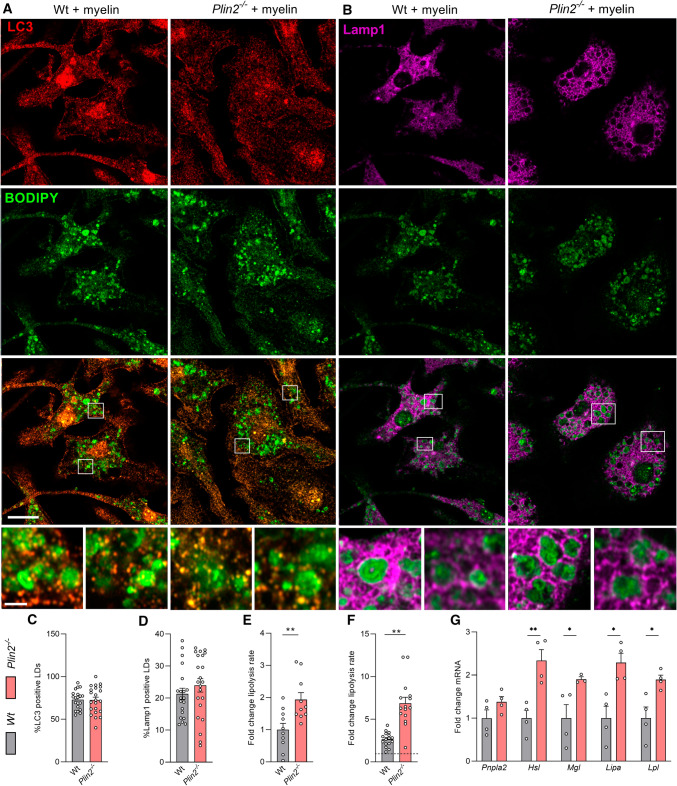
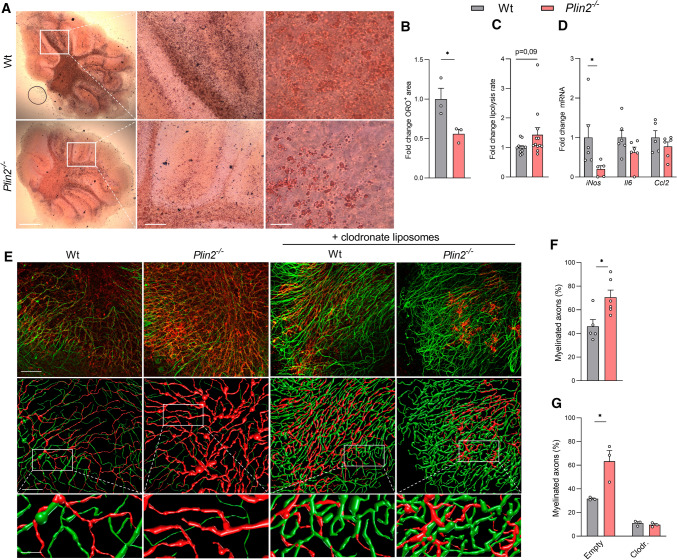
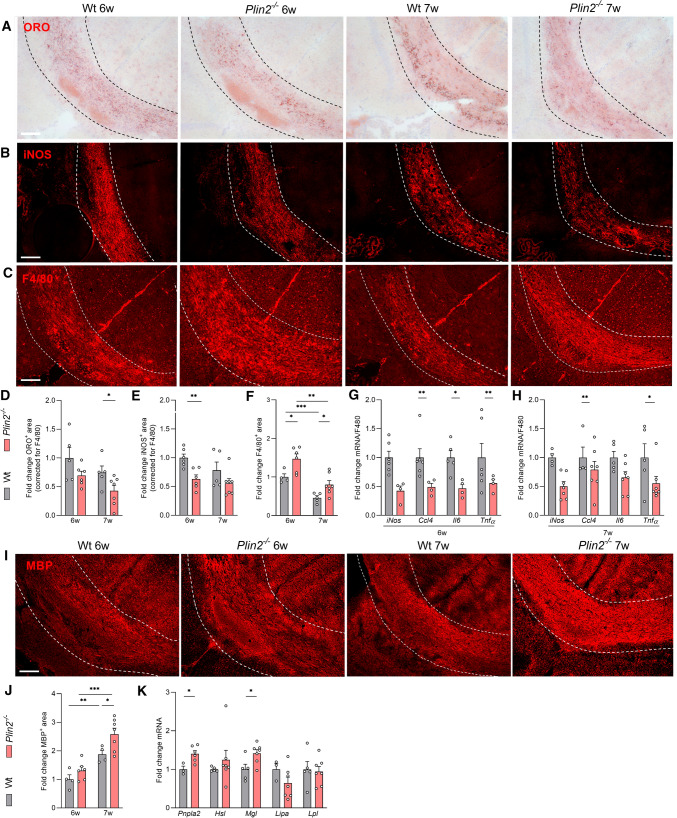
Foamy macrophages and microglia containing lipid droplets (LDs) are a pathological hallmark of demyelinating disorders affecting the central nervous system (CNS). We and others showed that excessive accumulation of intracellular lipids drives these phagocytes towards a more inflammatory phenotype, thereby limiting CNS repair. To date, however, the mechanisms underlying LD biogenesis and breakdown in lipid-engorged phagocytes in the CNS, as well as their impact on foamy phagocyte biology and lesion progression, remain poorly understood. Here, we provide evidence that LD-associated protein perilipin-2 (PLIN2) controls LD metabolism in myelin-containing phagocytes. We show that PLIN2 protects LDs from lipolysis-mediated degradation, thereby impairing intracellular processing of myelin-derived lipids in phagocytes. Accordingly, loss of Plin2 stimulates LD turnover in foamy phagocytes, driving them towards a less inflammatory phenotype. Importantly, Plin2-deficiency markedly improves remyelination in the ex vivo brain slice model and in the in vivo cuprizone-induced demyelination model. In summary, we identify PLIN2 as a novel therapeutic target to prevent the pathogenic accumulation of LDs in foamy phagocytes and to stimulate remyelination.
Keywords: Foamy macrophages; Inflammation; Lipid droplet associated protein; Lipolysis; Lipophagy.
© 2022. The Author(s), under exclusive licence to Springer Nature Switzerland AG.
Conflict of interest statement
The authors declare no competing interests exist.
Figures


References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
